14.5.2. Müllerian mimicry
In a contrasting set of relationships, called Müllerian mimicry, the model(s) and mimic(s) are all distasteful and warningly colored and all benefit from coexistence, as observers learn from tasting any individual. Unlike Batesian mimicry, in which the system is predicted to fail as the mimic becomes relatively more abundant, Müllerian mimicry systems gain through enhanced predator learning when the density of component distasteful species increases. “Mimicry rings” of species may develop, in which organisms from distant families, and even orders, acquire similar aposematic patterns, although the source of protection varies greatly. In the species involved, the warning signal of the co-models differs markedly from that of their close relatives, which are non-mimetic.
Interpretation of mimicry may be difficult, particularly in distinguishing protected from unprotected mimetic components. For example, a century after discovery of one of the seemingly strongest examples of Batesian mimicry, the classical interpretation seems flawed. The system involves two North American danaine butterflies, Danaus plexippus, the monarch or wanderer, and D. gilippus, the queen, which are chemically defended models each of which is mimicked by a morph of the nymphaline viceroy butterfly (Limenitis archippus) (Fig. 14.8). Historically, larval food plants and taxonomic affiliation suggested that viceroys were palatable, and therefore Batesian mimics. This interpretation was overturned by experiments in which isolated butterfly abdomens were fed to natural predators (wild-caught redwing blackbirds). The possibility that feeding by birds might be affected by previous exposure to aposematism was excluded by removal of the aposematically patterned butterfly wings. Viceroys were found to be at least as unpalatable as monarchs, with queens least unpalatable. At least in the Florida populations and with this particular predator, the system is interpreted now as Müllerian, either with the viceroy as model, or with the viceroy and monarch acting as co-models, and the queen being a less well chemically protected member that benefits through the asymmetry of its palatability relative to the others. Few such appropriate experiments to assess palatability, using natural predators and avoiding problems of previous learning by the predator, have been reported and clearly more are required.
If all members of a Müllerian mimicry complex are aposematic and distasteful, then it can be argued that an observer (predator) is not deceived by any member — and this can be seen more as shared aposematism than mimicry. More likely, as seen above, distastefulness is unevenly distributed, in which case some specialist observers may find the least well-defended part of the complex to be edible. Such ideas suggest that true Müllerian mimicry may be rare and/or dynamic and represents one end of a spectrum of interactions.
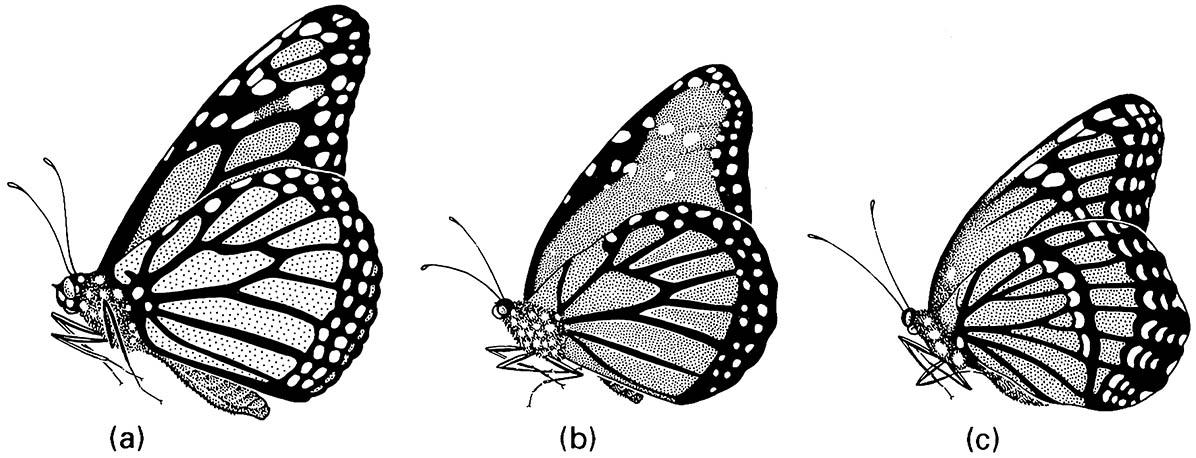
(a) the monarch or wanderer (Danaus plexippus); (b) the queen (D. gilippus); (c) the viceroy (Limenitis archippus). (After Brower 1958)

