Box 16.1. Bemisia tabaci biotype B: a new pest or an old one transformed?
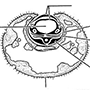
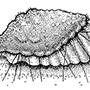
Bemisia tabaci, often called the tobacco or sweetpotato whitefly, is a polyphagous and predominantly tropical—subtropical whitefly (Hemiptera: Aleyrodidae) that feeds on numerous fiber (particularly cotton), food, and ornamental plants. Nymphs suck phloem sap from minor veins (as illustrated diagrammatically on the left of the figure, after Cohen et al. 1998). Their thread-like mouthparts (section 11.2.3; Fig. 11.4) must contact a suitable vascular bundle in order for the insects to feed successfully. The whiteflies cause plant damage by inducing physiological changes in some hosts, such as irregular ripening in tomato and silverleafing in squash and zucchini (courgettes), by fouling with excreted honeydew and subsequent sooty mold growth, and by the trans- mission of more than 70 viruses, particularly geminiviruses (Geminiviridae).
Infestations of B. tabaci have increased in severity since the early 1980s owing to intensive continuous cropping with heavy reliance on insecticides and the possibly related spread of what is either a virulent form of the insect or a morphologically indistinguishable sibling species. The likely area of origin of this pest, often called B. tabaci biotype B, is the Middle East, perhaps Israel. Certain entomologists (especially in the USA) recognize the severe pest as a separate species, B. argentifolii, the silverleaf whitefly (the fourth-instar nymph or “puparium” is depicted on the right, after Bellows et al. 1994), so-named because of the leaf symptoms it causes in squash and zucchini. B. argentifolii exhibits minor and labile cuticular differences from the true B. tabaci (often called biotype A) but comparisons extended to morphologies of eight biotypes of B. tabaci found no reliable features to separate them. However, clear allozyme, nuclear, and mitochondrial genetic information allows separation of the non-B bio-types of B. tabaci. Nucleotide sequences of the 18S rDNAs of biotypes A and B and the 16S rDNAs of their bacterial endosymbionts are essentially identical, suggesting that these two whiteflies are either the same or very recently evolved species. Some biotypes show variable reproductive incompatibility, as shown by crossing experiments, which may be due to the presence of strain- or sex-specific bacteria, resembling the Wolbachia and similar endosymbiont activities observed in other insects (section 5.10.4). Populations of B. tabaci biotype A are eliminated wherever biotype B is introduced, suggesting that incompatibility might be mediated by microorganisms. Indeed, the bacterial faunas of B. tabaci biotypes A and B show some differences in composition, consistent with the hypothesis that symbiont variation may be associated with biotype formation. For example, recently it was shown that biotype A, but not biotype B, is infected by a chlamydia species (Simkaniaceae: Fritschea bemisiae) and it is possible that the presence of this bacterium influences the fitness of its host whitefly. Furthermore, endosymbionts in some other Hemiptera have been associated with enhanced virus transmission (section 3.6.5), and it is possible that endosymbionts mediate the transmission of geminiviruses by B. tabaci biotypes.
The sudden appearance and spread of this apparently new pest, B. tabaci biotype B, highlights the importance of recognizing fine taxonomic and biological differences among economically significant insect taxa. This requires an experimental approach, including hybridization studies with and without bacterial associates. It is probable that B. tabaci is a sibling species complex, in which most of the species currently are called biotypes, but some forms (e.g. biotypes A and B) may be conspecific although biologically differentiated by endosymbiont manipulation. In addition, it is feasible that strong selection, resulting from heavy insecticide use, may select for particular strains of whitefly or bacterial symbionts that are more resistant to the chemicals.
Effective biological control of Bemisia whiteflies is possible using host-specific parasitoid wasps, such as Encarsia and Eretmocerus species (Aphelinidae). However, the intensive and frequent application of broad- spectrum insecticides adversely affects biological control. Even B. tabaci biotype B can be controlled if insecticide use is reduced.
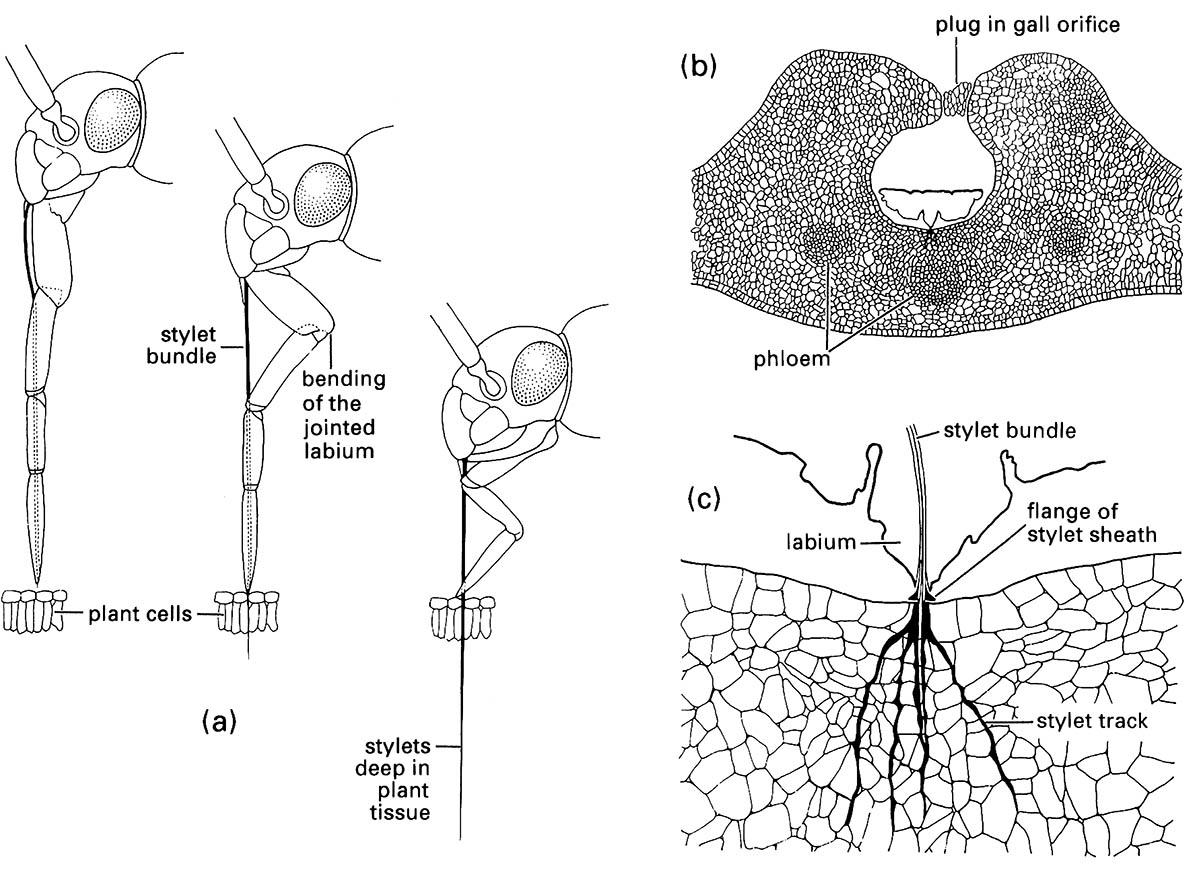
(a) penetration of plant tissue by a mirid bug showing bending of the labium as the stylets enter the plant; (b) transverse section through a eucalypt leaf gall containing a feeding nymph of a scale insect, Apiomorpha (Eriococcidae); (c) enlargement of the feeding site of (b) showing multiple stylet tracks (formed of solidifying saliva) resulting from probing of the parenchyma. ((a) After Poisson 1951)