Слуховая коммуникация
Насекомые производят множество разнообразных звуков и в большинстве случаев на какой-то стадии развития именно с помощью звуков осуществляется связь между особями. У некоторых насекомых звук создаётся в процессе их нормальной жизнедеятельности без помощи каких-либо специальных звукоиздающих структур. Наиболее известный пример такого рода — это жужжание летящего или парящего насекомого. Жужжание создаётся в результате очень быстрой вибрации крыльев и склеритов груди. У некоторых насекомых, например у комаров, звуки, возникающие от биения крыльев летящей самки, вызывают у самца реакцию спаривания. У других насекомых такие звуки не служат средством коммуникации.
Исходя из особенностей человеческого слуха, трудно оценить качество звуков, производимых насекомыми. Продолжительный звук, например звук от вибрации крыльев, менее обычен, чем дискретные звуковые пульсы, разделенные периодами тишины. Насекомые способны улавливать более широкий диапазон частот, особенно в области высоких частот, чем человеческое ухо, воспринимающее звуки от 60 до 16 000 циклов в секунду (ц/с). Люди не слышат биения крыльев бабочек (отряд Lepidoptera), частота которого около 20 ц/с. Частота биения крыльев пчелы Apis составляет около 250 ц/с. Шум летящей саранчи Schistocerca создаётся звуком с частотой от 60 до 6 400 ц/с, но в основном в диапазоне 3 200 — 5 000 ц/с. Частота звука, создаваемая биением головы термита (Isoptera) по древесине, около 1 000 ц/с. У различных видов сверчков (Orthoptera) в результате трения крыльев (элитральная стридуляция) создаются звуки с большим диапазоном частот — от 2 000 — 10 000 ц/с до очень высоких, порядка 50 000 — 100 000 ц/с. Стрекотание сверчка имеет характерную для него постоянную частоту и амплитуду и представляет собой очень чистый звук. Частота звука, создаваемого тимбалами цикад (Hemiptera), равна примерно 4 500 ц/с.
Помимо звуков, издаваемых при проявлении нормальной жизнедеятельности насекомого, существуют другие, производимые иными способами:
- ударение какой-либо частью тела по субстрату;
- фрикционное трение двух частей тела друг о друга (обычно называется стридуляцией);
- вибрация мембран;
- выталкивание струй воздуха.
Среди множества видов насекомых встречаются многочисленные вариации этих способов.
1. При издавании звуков посредством удара о субстрат насекомые пользуются различными частями тела. Кобылка Oedipoda ударяет по поверхности субстрата задней голенью. Самец делает примерно 12 ударов в секунду, самка несколько меньше. Жук Xestobium стучит по субстрату головой и производит своеобразную «очередь» хлопков. Солдаты некоторых термитов совершают колебательные движения, и при этом голова и кончики мандибул ударяют по древесине так, что получается серия звуков, которую воспринимают другие обитатели термитника (рабочие и личинки). Другие касты термитов также производят звуки при ударе головой по древесине, но с более низкой интенсивностью.
2. Звуки, возникающие от трения различных частей тела друг о друга, известны у многих насекомых. Этот способ хорошо развит в отрядах Orthoptera, Неteroptera и Coleoptera. У некоторых прямокрылых передний край заднего крыла ударяет по утолщённым жилкам переднего, вызывая вибрацию последнего; в результате такой вибрации и возникают трескучие звуки. У некоторых саранчовых на внутренней поверхности заднего бедра имеется ряд маленьких зубчиков (рис. 6.17), которые трутся о переднее крыло, производя таким образом звук. У различных сверчков на одной из элитр имеется ряд зубчиков (щётка), а на другой — гребень (скребок). При соприкосновении элитр зубчики трутся о гребень, элитры вибрируют, и возникает звук (рис. 6.18). У других насекомых, например у жуков, гребень и щётка расположены соответственно на ноге и на теле (рис. 6.19). Для генерации звуков жуки используют самые различные части своего склеротизированного экзоскелета, но особенно элитры.
3. Вибрирующие мембраны, или тимбальный механизм, обнаружен у некоторых равнокрылых, в частности у цикад, у других полужесткокрылых и у бабочек. Наиболее интересное приспособление развилось у цикад (рис. 6.20). У этих насекомых у основания брюшка имеются карманы, или полости, в которых располагается несколько мембран.
Внутренняя поверхность одной из этих мембран соединена с мышечным волокном. Сокращение этого волокна втягивает мембрану внутрь, а расслабление мышцы позволяет ей вернуться в исходное положение. Эти движения мембраны происходят с большой скоростью и создают звуковые волны. Остальные мембраны функционируют в качестве рефлекторов звука.
4. Ещё одним способом образования звука у насекомых является проталкивание воздуха через или над маленьким отверстием. У бражника Acherontia atropos звук возникает в результате втягивания и выталкивания воздуха через хоботок и создания воздушных струй над эпифаринксом (надглоточником).
Звуки могут служить сигналами, воспринимаемыми другими видами (межвидовые сигналы) или другими особями того же вида (внутривидовые). Неупорядоченные звуки без регулярно повторяющихся пульсов служат, по-видимому, для защиты или предупреждения и могут быть как меж-, так и внутривидовыми. Очевидно, к такому типу звуков относится стридуляция у самок и самцов некоторых Heteroptera, жуков и куколок бабочек. Эти реакции тревоги сопровождаются иногда другими защитными реакциями, такими, как шипение, раздающееся в тот момент, когда бабочка павлиноглазка раскрывает крылья, чтобы продемонстрировать яркие глазчатые пятна.
Упорядоченные звуки, повторяющиеся с определённой частотой, наиболее характерны для внутривидовых коммуникаций. Как правило, это связано с процессом ухаживания. У Orthoptera различают пять типов звуковых сигналов, означающих призыв, ухаживание, копуляцию, нападение и тревогу. Каждый из этих типов отличается по частоте и характеру пульсов. У ряда насекомых, например цикад, определённые звуки вызывают агрегацию особей. Самцы и самки одного вида собираются в каком-то определённом месте, например в кроне тех или иных деревьев. Пение некоторых цикад обеспечивает половую изоляцию видов. В Северной Америке три вида рода Magicicada занимают одно и то же местообитание. Однако особи каждого вида идентифицируют особей своего и чужого видов по пению. Кроме того, разные виды поют хором в разное время дня.
У полуобщественных и общественных насекомых звуки служат средством различения особей своего вида и своей колонии. Шершни производят в своих гнёздах целый ряд различных звуков-сигналов. Роение пчёл отчасти стимулируется продолжительным жужжанием, производимым несколькими рабочими особями, возбуждённо шныряющими по улью.
У общественных насекомых звуки служат также сигналами для поисков пищи и постройки гнезда. У общественных насекомых звук является важнейшим средством коммуникации, однако он не исключает других средств коммуникации, например химических или тактильных. Взаимодействие и интеграция различных типов коммуникации хорошо проявляются у классического общественного насекомого — медоносной пчелы Apis mellifera. В танце пчёл, считавшемся когда-то проявлением зрительной коммуникации, обнаружен целый комплекс сигналов, включающих и звук, и проприоцепцию. Какой из типов коммуникации играет ведущую роль в танце, до сих пор не установлено. Танец представляет собой ряд специфических стереотипных движений, совершаемых пчелой — сборщицей внутри улья на вертикальной поверхности сот. Пчела-сборщица, отыскавшая источник корма, приносит в улей пыльцу и нектар и затем снова вылетает к этому источнику. Возвращаясь второй раз, она совершает танец в месте скопления большого числа рабочих пчёл, причём характер танца меняется в зависимости от расстояния до источника пищи. Если он находится близко от улья, то выполняется круговой танец (рис. 6.21, А). Пчела-сборщица движется по кругу, меняя направление движения.
Считается, что такой танец означает лишь только то, что источник пищи недалеко, но он не несёт информации о направлении полёта и расстоянии до него. Если источник пищи находится на среднем удалении от улья, то выполняется серпообразный танец (открытая «восьмёрка»), при этом круговой танец постепенно переходит в движение по открытой «восьмёрке». В том случае, если пища далеко, выполняется виляющий танец (рис. 6.21, Б). В этом танце пчела, двигаясь по фигуре в виде открытой «восьмёрки», совершает быстрые виляющие движения брюшком во время прямой части своего пробега. Полагают, что именно эта часть танца даёт информацию о расстоянии до источника корма и направлении к нему. Жужжание, производимое пчелой-сборщицей во время пробега по прямой, его ритм и суммарная продолжительность звукового сигнала, возможно, используются другими пчёлами в улье для установления расстояния до источника корма.
Вероятно, что эта информация дополняется информацией, представленной числом виляний брюшка во время прямого пробега. Угол между направлением прямого пробега пчелы-сборщицы и проведённой к этой линии вертикалью соответствует углу между двумя линиями, проведёнными от улья к солнцу и к источнику корма (рис. 6.22). Пробег по прямой, направленной вертикально вверх, указывает на то, что источник пищи находится в том же направлении, что и солнце; если прямая направлена вертикально вниз, то, следовательно, источник корма находится в направлении, противоположном солнцу.
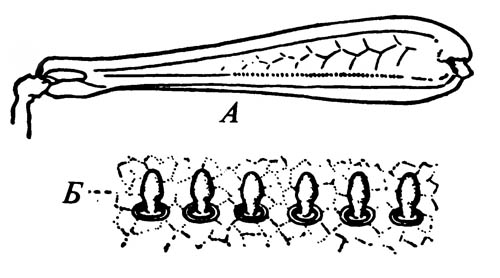
6.17. Пилочка на внутренней поверхности бедра задней ноги сверчка
A. Заднее бедро Stenobothrus. Б. Пилочка, сильно увеличено.
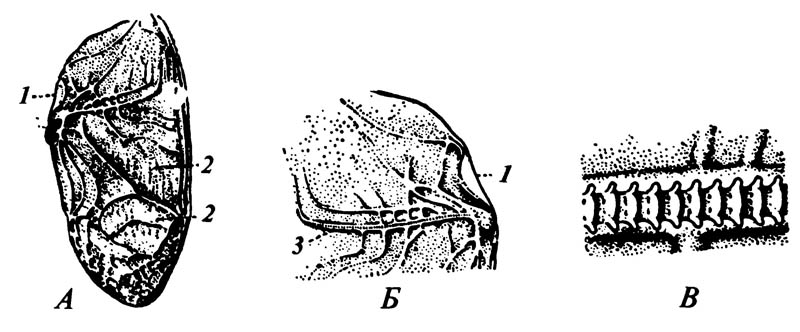
6.18. Переднее крыло сверчка Gryllus с пилочкой и скребком
А. Вид сверху; часть сложенного крыла, прилегающая к брюшку, не видна. Б. Основание крыла, вид снизу. В. Пилочка, сильно увеличено. 1 — скребок; 2 — мембрана; 3 — пилочка.

6.19. Стридуляционный орган личинки Pussalus
1, 2 — части заднегруди; 3 — тазик второй
4 — пилочка; 5 — базальная часть бедра передней ноги; 6 — волоски с хитиновыми выростами у их основания; 7 — уменьшенная задняя нога, превращённая в скребок
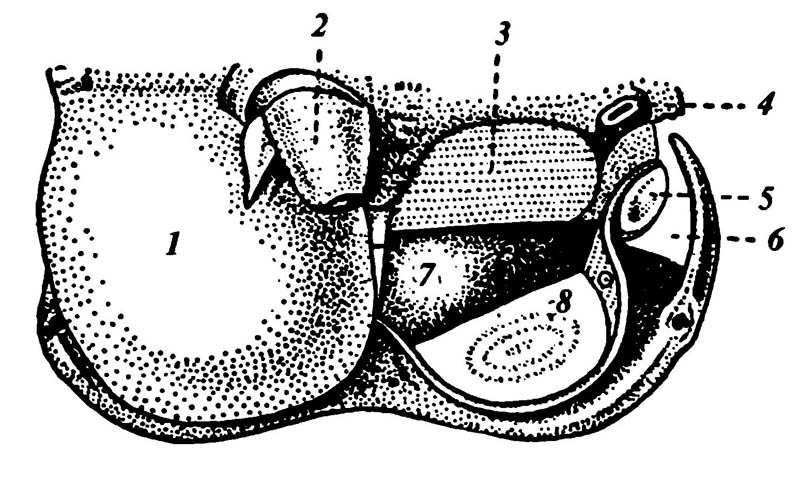
6.20. Музыкальный аппарат цикады
1 — крышечка (на другой стороне крышечка удалена); 2 — основание ноги, 3 — складчатая мембрана, 4 — дыхальце; 5 — вибрирующая мембрана (тимбал); 6 — латеральная полость; 7 — вентральная полость; 8 — зеркальце.
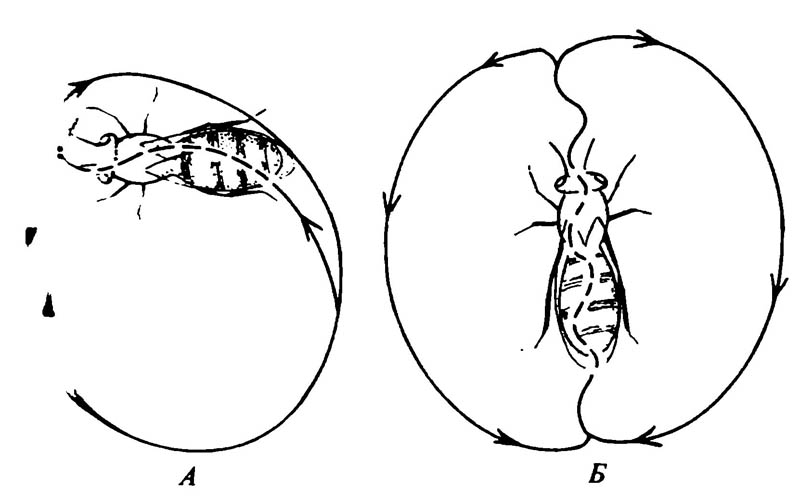
6.21. Танцы медоносной пчелы Apis mellifera
А. Круговой танец пчелы-сборщицы, информирующий о близости источника корма к улью. Б. «Виляющий» танец пчелы-сборщицы, информирующий об удалённости источника корма от улья
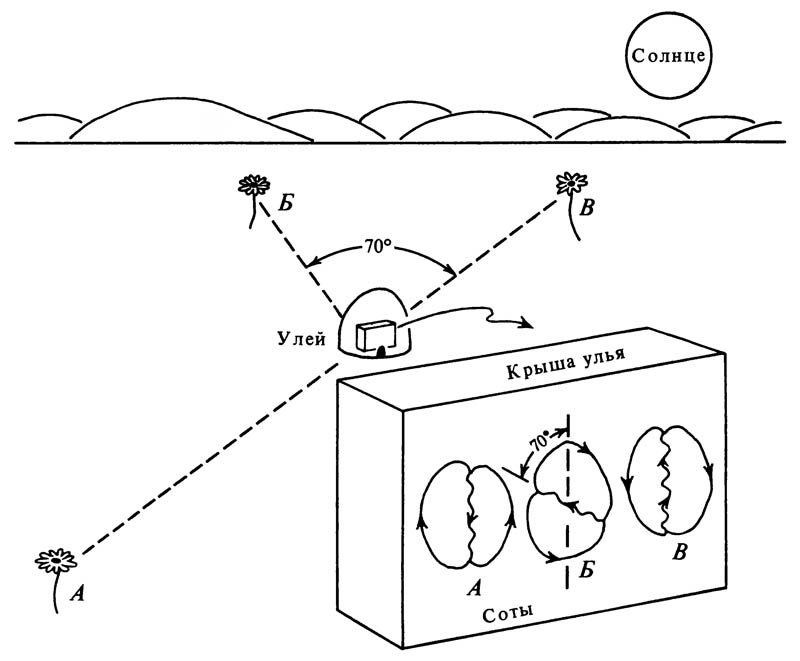
6.22. Направление, в котором расположен удалённый источник корма, показывается «виляющим» танцем медоносной пчелы
Угол между направлением пробега с «вилянием» и вертикалью равен, по-видимому, углу между направлениями к солнцу и к источнику корма, спроектированному на горизонтальную плоскость

Зрительная коммуникация


