Box 14.1. Avian predators as selective agents for insects
Henry Bates, who was first to propose a theory for mimicry, suggested that natural enemies such as birds selected among different prey such as butterflies, based upon an association between mimetic patterns and unpalatability. A century later, Henry Kettlewell argued that selective predation by birds on the peppered moth (Geometridae: Biston betularia) altered the proportions of dark- and light-colored morphs (Fig. 14.2) according to their concealment (crypsis) on natural and industrially darkened trees upon which the moths rested by day. Amateur lepidopterists recorded that the proportion of the dark (“melanic”) carbonaria form dramatically increased as industrial pollution in- creased in northern England from the mid-19th century. Elimination of pale lichen on tree trunk resting areas was suggested to have made normal pale morphs more visible against the sooty, lichen-denuded trunks (as shown in Fig. 14.2b), and hence they were more susceptible to visual recognition by bird predators. This phenomenon, termed industrial melanism, often has been cited as a classic example of evolution through natural selection.
The peppered moth/avian predation story has been challenged for its experimental design and procedures, and biased interpretation. The case depended upon:
- birds being the major predators rather than night-flying, pattern-insensitive bats;
- moths resting “exposed” on trunks rather than under branches or in the canopy;
- dark and pale morphs favoring the cryptic background appropriate to their patterning;
- crypsis to the human eye being quantifiable and equating to that for moth-feeding birds;
- selection being concentrated in the adult stage of the moth’s life cycle;
- genes responsible for origination of melanism acting in a particular way, and with very high levels of selection. None of these components have been confirmed. Evolution undoubtedly has taken place. The proportions of dark morphs (alleles for melanism) have changed through time, increasing with industrialization, and reducing as “post-industrial” air quality improves. However, the centrality of avian predation acting as a force for natural selection in B. betularia is no longer so evident.
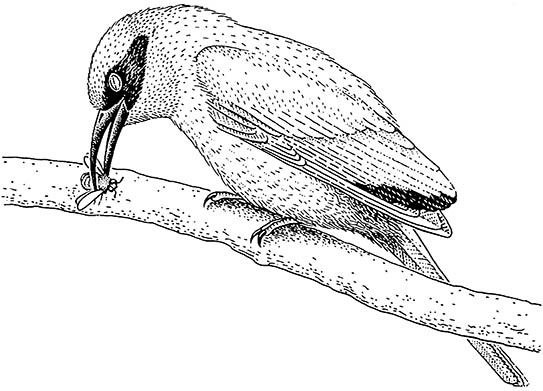
More convincing is the demonstration of directly observed predation, and inference from beak pecks on the wings of butterflies and from experiments with color-manipulated daytime-flying moths. Thus, winter- roosting monarch butterflies (Danaus plexippus) are fed upon by black-backed orioles (Icteridae), which browse selectively on poorly defended individuals, and by black-headed grosbeaks (Fringillidae), which appear to be completely insensitive to the toxins. Specialized predators such as Old World bee-eaters (Meropidae) and neotropical jacamars (Galbulidae) can deal with the stings of hymenopterans (the red-throated bee-eater, Merops bullocki, is shown here destinging a bee on a branch, after Fry et al. 1992) and the toxins of butter- flies, respectively. A similar suite of birds selectively feeds on noxious ants. The ability of these specialist predators to distinguish between varying pattern and edibility may make them selective agents in the evolution and maintenance of defensive mimicry.
Birds are observable insectivores for laboratory studies: their readily recognizable behavioral responses to unpalatable foods include head-shaking, disgorging of food, tongue-extending, bill-wiping, gagging, squawking, and ultimately vomiting. For many birds, a single learning trial with noxious (Class I) chemicals appears to lead to long-term aversion to the particular insect, even with a substantial delay between feeding and illness. However, manipulative studies of bird diets are complicated by their fear of novelty (neophobia), which, for example, can lead to rejection of prey with startling and frightening displays (section 13.2). Conversely, birds rapidly learn preferred items, as in Kettlewell’s experiments in which birds quickly recognized both Biston betularia morphs on tree trunks in his artificial set-up.
Perhaps no insect has completely escaped the attentions of predators and some birds can overcome even severe insect defenses. For example, the lubber grasshopper (Acrididae: Romalea guttata) is large, gregarious, and aposematic, and if attacked it squirts volatile, pungent chemicals accompanied by a hissing noise. The lubber is extremely toxic and is avoided by all lizards and birds except one, the loggerhead shrike (Laniidae: Lanius ludovicanus), which snatches its prey, including lubbers, and impales them “decoratively” upon spikes with minimal handling time. These impaled items serve both as food stores and in sexual or territorial displays. Romalea, which are emetic to shrikes when fresh, become edible after two days of lardering, presumably by denaturation of the toxins. The impaling behavior shown by most species of shrikes thus is preadaptive in permitting the loggerhead to feed upon an extremely well-defended insect. No matter how good the protection, there is no such thing as total defense in the arms race between prey and predator.
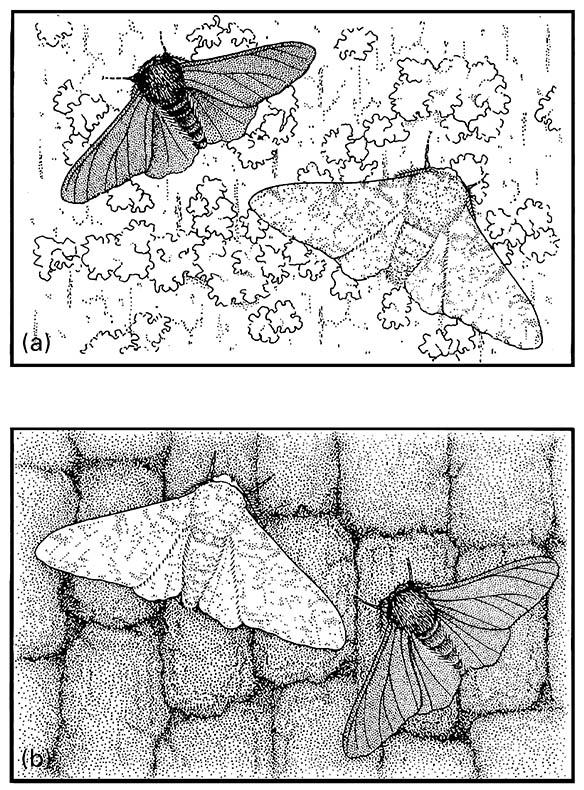
(a) pale, lichen-covered; and (b) dark trunks.



