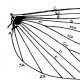
Поперечные жилки
В общем поперечные жилки принято называть по тем продольным жилкам, между которыми они лежат. Так, поперечные жилки, связывающие радиальную жилку с передней ветвью сектора, получат в этом случае наименования 1r-s, 2r-s и так далее; связывающие заднюю ветвь медиальной жилки с кубитальной будут обозначены 1m-cu, 2m-cu и так далее.
Поперечные жилки между ветвями одной и той же продольной жилки часто имеют особые названия в разных систематических группах. Существует и несколько общепринятых исключений. Поперечная жилка между костальной и субкостальной жилками в основании крыла называется плечевой (h), а остальные, лежащие между костальной и следующей за ней жилкой, — просто коcтальными поперечными жилками.
Происхождение. Существует несколько теорий происхождения такого уникального органа, каковым является крыло насекомых. Поскольку оба наиболее примитивных отряда крылатых насекомых, Ephemeroptera и Odonata, а также другой весьма примитивный отряд, Р1еcoptera, имеют водных нимф, вначале предполагали, что крылья возникли как плавательные лопасти, которые помогали личинкам продвигаться в толще воды. Однако трудности, возникающие при попытке реконструкции форм, переходных между плавательной лопастью и возможным крылом, заставляют признать эту теорию несостоятельной. Согласно другой теории, предложенной Александером и Броуном (Alexander, Brown, 1963), крылья возникли из боковых выростов груди, связанных с половой активностью.
До настоящего времени каких-либо половых структур, которые могли бы послужить предшественниками органов полёта, не было обнаружено ни на груди ископаемых насекомых, ни у современных представителей. Наиболее достоверной представляется гипотеза, в соответствии с которой крылья берут начало от боковых лопастевидных выростов спинки груди; возникли они, возможно, в качестве защитных образований или в результате уплощения тела как адаптации к проникновению в щели, трещины или под кору. Если насекомое было чем-то обеспокоено или встревожено, эти выступающие части помогали ему спланировать, не потеряв правильной ориентации тела, чтобы быть готовым к бегству после падения с высоты вниз. Однако даже и в этом случае необходимо было особое стечение обстоятельств.
Насекомые должны были часто посещать поверхности, например, листья, стволы деревьев, отвесные скалы), находившиеся на расстоянии нескольких метров от уровня почвы: в противном случае при падении они были бы не в состоянии набрать скорость, достаточную для того, чтобы боковые выросты спинки смогли взаимодействовать с воздушным потоком. Флауер (Flower, 1964) подсчитал, что коротконогие насекомые с длиной тела немного менее 1 см и лишь со слабой пластинкой вдоль краёв груди вполне могли удовлетворять этим требованиям и в конце падения достигать устойчивого равномерного скольжения вниз. Так как насекомыми питались прежде всего быстро бегающие хищники, такие, как пауки и многоножки, скорость и способность маневрировать в воздухе давали бы им преимущества для выживания.
Среди современных насекомых подобным аэродинамическим требованиям удовлетворила бы, например, чешуйница Thermobia. Это и некоторые другие примитивные бескрылые насекомые, такие, как Machilis, имеют и иные приспособления, которые могли бы оказаться полезными при попытках подобного планирующего бегства, грудь расположена вблизи центра тяжести тела, а три пары больших сильных грудных ног могут обеспечить сильный толчок для поспешного спасения. Теперь представляется вполне вероятным, что способность к планированию у организмов, напоминающих нынешних Thermobia, явилась предпосылкой для развития полёта.
Другой факт, который следует принять во внимание при выяснении путей эволюции полёта насекомых, — это наличие на переднеспинке многих древнейших ископаемых крылатых насекомых плоских боковых выростов, названных паранотальными лопастями и служащих, вероятно, для планирования. Кроме того, эти лопасти имели набор жилок, по-видимому гомологичный главным жилкам типичного подвижно сочленённого функционирующего крыла. Эти дополнительные сведения позволяют восстановить ход эволюции крыла насекомых с высокой степенью достоверности.
Когда было достигнуто устойчивое скольжение вниз, начало играть свою роль давление отбора в направлении расширения спинных выростов и превращения их в лопасти или плоскости, которые позволяли бы сократить время до начала скольжения и увеличить протяжённость самого скольжения. Вначале лопасть походила, возможно, на изображённую на рисунке А, затем она увеличивалась, проходя через стадии, показанные на рисунке Б, В, Г и Д. Сходные лопасти изначально имелись, по-видимому, на всех трёх грудных сегментах. По мере увеличения размеров лопастей в них проникали ответвления трахей; у взрослых особей они превращались в места утолщения покровов крыла, то есть в жилки. На рисунке Е изображена наиболее крупная из известных нам лопастей, обнаруженных на переднегруди. Возможно, что на этой стадии в основании крыла уже был сформировавшийся шарнир, и с этого времени средне- и заднегрудные крылья становились больше, а лопасти переднегруди уменьшались.
Первые маленькие крылья были, видимо, ещё довольно толстыми и неуклюжими. По мере увеличения крыла его жилки становились более мощными, а крыловая мембрана между ними — более тонкой и лёгкой. Наиболее древнее из известных действующих крыльев было многократно сложено в продольном направлении подобно вееру, причём жилки шли по местам перегибов, кроме того, имелась сеть поперечных жилок, связывавших между собой продольные. Такая комбинация вееро-образной или складчатой структуры и запоминающего арматурные прутья каркаса позволяла прочной, но тонкой мембране быть достаточно жёсткой для полёта.
Последующей стадией эволюции крыла была перегруппировка аксиллярных склеритов и пластинок, сделавшая возможным диагональное складывание крыла у его основания. При такой организации крылья могли складываться вдоль анальной и базальной складок назад и плашмя над телом. На том же этапе эволюции большая часть складок крыла исчезла и область между R и Cu стала плоской.
В разных отрядах насекомых произошли разнообразные изменения в жилковании крыльев. У одних появились добавочные продольные жилки, у других различные жилки либо сливаются, либо отсутствуют. То же относится и к поперечным жилкам. У крупных насекомых, характеризующихся быстрым полётом, обычно имеются длинные и узкие крылья ; порхающий полет, как у дневных бабочек, коррелирует с более короткими и широкими крыльями. У мелких насекомых, например тлей, жилкование может быть в значительной степени редуцировано, а полет часто слабый, почти пассивный. Каждый тип модификации крыла находится в связи с поведением и образом жизни взрослого насекомого.
Во многих группах способность к полёту полностью утрачена, как, например, у прямокрылых семейства Rhaphidophoridae или блох, которые лишены крыльев.
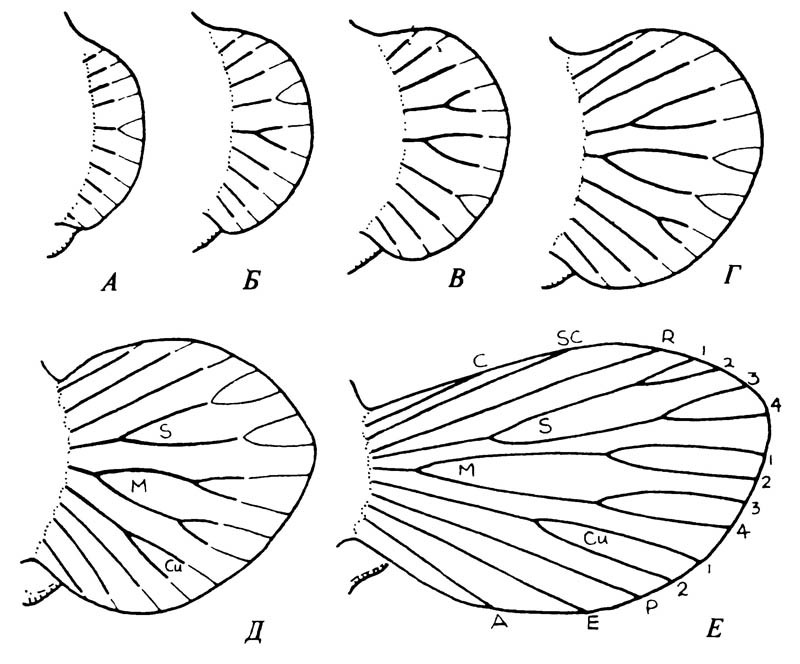
Гипотетические стадии развития жилкования паранотальных лопастей (А — Д), приводящие к жилкованию примитивного крыла (Е)

Форма крыла Paleoptera на поперечном срезе. Сильная гофрировка крыловой пластинки за счёт чередования выступающих и погруженных жилок придаёт крылу дополнительную жёсткость
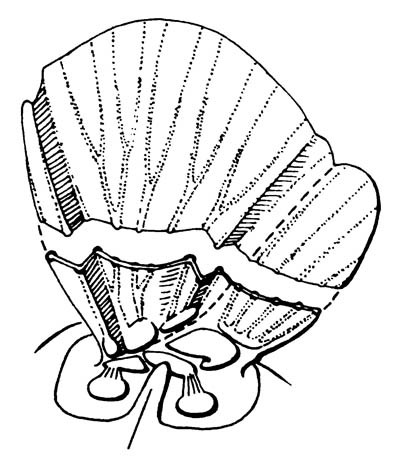
Форма крыла Neoptera на поперечном срезе. Гофрировка крыловой пластинки изменилась, позволив крылу складываться, не проигрывая при этом в жёсткости