Предки членистоногих
Нет сомнения, что Arthropoda произошли от червеподобного предка, близкого по общему строению к Annelida, или кольчатым червям. Тело этого предка (рис. 2.1) состояло из серии одинаковых сегментов, каждый из которых представлял собой замкнутое кольцо. Голова имела простое строение и, по-видимому, несла чувствительные щетинки. Ротовое отверстие было расположено на вентральной стороне тела между головой и первым сегментом тела. Из-за своего положения впереди ротового, или стомодеального, отверстия головной отдел этих животных называют простомиумом.
Как отмечал Мэнтон (Manton, 1973), этот червеподобный предок членистоногих отличался от современных Annelida по ряду важных черт в строении тела. У Annelida каждый сегмент тела имеет свою полость, или целом, причём целомы разных сегментов разделены перегородками — септами. В каждом сегменте имеются выделительные органы (парные нефридии), открывающиеся наружу выводными протоками. Ноги, если они имеются, также расположены посегментно и характеризуются примитивным строением. У гипотетических членов ряда, ведущего к Arthropoda, отсутствовали межсегментные перегородки и полость тела простиралась по всей его длине. Выделительная система представляла собой один орган, обслуживавший весь организм животного. Ноги, если они имелись, были с более специализированной, чем у Annelida, мускулатурой. На рис. 2.1 показаны ступени эволюционного развития гипотетического предка от простой преартроподной организации через генерализованный артроподный тип строения к насекомым.
Первая важная ступень состояла в образовании на вентральной стороне каждого сегмента пары конечностей, или ног, обеспечивавших передвижение животного (рис. 2.1). Очевидно, последний сегмент, перипрокт, на котором располагалось анальное отверстие, никогда не имел конечностей.
Следующим вероятным изменением было слияние конечностей первого сегмента и образование лабрума — вентрального придатка, функция которого состояла в направлении пищи к ротовому отверстию. Конечности второго сегмента переместились на дорсальную сторону и превратились в антенны. Конечности третьего сегмента, имеющиеся на ранних эмбриональных стадиях насекомых, исчезают как самостоятельные структуры на более поздних стадиях развития. Один за другим эти первые три сегмента, носящие соответственно названия лабральный, антеннальный и интеркалярный, сливаются с анцестральным простомиумом, образуя сложную голову, составленную из исходных шести сегментов. Ганглии этих сегментов сливаются с первичным мозгом, или архицеребрумом, и образуется сложный головной мозг членистоногих.
Эти изменения в строении головы и головного мозга сопровождались другим эволюционным преобразованием, важным для питания животных. Ротовое отверстие, расположенное вначале позади лабрального сегмента, затем переместилось в положение позади антеннального сегмента и, наконец, оказалось позади интеркалярного сегмента. Несомненно, что на этом доартроподном этапе эволюции конечности нескольких следующих сегментов, пока лишь слегка модифицированные, служили для направления пищи к ротовому отверстию. Ротовое отверстие, смещённое кзади, вместе с лабрумом, также переместившимся назад, образовало нечто вроде кармана, где пища могла накапливаться до того, как животное проглатывало её. Поскольку термин «простомиум» не соответствовал более тому строению, которое приобрела первичная голова после перемещения ротового отверстия назад, она получила название акрон.
Перемещение ротового отверстия из положения между акроном и сегментом 1 в положение между сегментами 3 и 4 требует некоторого разъяснения. На ранних стадиях эмбрионального развития структуры вентральной части тела формировались до того, как сегменты приобретали свою окончательную форму-форму колец. Предшественниками будущих нервных центров, или ганглиев, были расположенные в каждом сегменте парные, но не связанные между собой латеральные массы нервной ткани, называемые нейромерами (нервными телами). На ранних стадиях эмбриогенеза эти отдельные нейромеры, по-видимому, продвигались вперёд по обеим сторонам от ротового отверстия и затем сливались с первичным мозгом.
Примерно на этой стадии эволюции членистоногих произошло превращение конечностей сегмента 4 в хватательные и режущие придатки, называемые мандибулами. Однако нейромеры этого сегмента не переместились вперёд и ротовое отверстие осталось между сегментами 3 и 4.
Эта стадия эволюции представлена тремя группами Onychophora, Pentastomida и Tardigrada, особенно чётко характеризующие её черты строения можно наблюдать у современных представителей Onychophora (Peripatus и близкие к нему формы, рис. 2.2.). Онихофоры имеют мягкое тело с посегментно расположенными парными конечностями, хотя внешние признаки сегментации чётко не выражены. Ноги онихофор, называемые лобоподиями, не сегментированы, но имеют специальные мышцы, прикреплённые к неподвижным основаниям (Manton, 1973). У них также имеется пара придатков, сходных по своему виду и положению с мандибулами членистоногих, однако гомология их пока не доказана. Даже если это и так, онихофор всё-таки следует рассматривать как отдельную чёткую ветвь этого направления эволюции.
Представители двух других групп, Tardigrada (рис. 2.3.) и Pentastomida (рис. 2.4.), претерпели столь сильную редукцию организации, что их родственные связи требуют дальнейшего тщательного изучения. Tardigrada (тихоходки, или водяные медведи; рис. 2.3.) — это крошечные животные, живущие во влажном мху, а также в пресной и солёной воде. Их тело, никогда не превышающее в длину 11 мм, имеет 4 пары бугорковидных ног, заканчивающихся коготками. На голове нет ни чётко различимых ротовых придатков, ни каких-либо других структур. Pentastomida (язычковые, или пятиустки; рис. 2.4.) — небольшая группа животных с червеподобными взрослыми формами и крошечными четырёхногими неполовозрелыми стадиями, по общему виду похожими на клещей. Пятиустки являются внутренними паразитами ряда позвоночных.
Следующая стадия эволюции той ветви, которая ведёт к членистоногим, характеризуется некоторыми изменениями в сторону специализации.
- Ноги приобрели членистое строение. что способствовало развитию локомоторной функции.
- Некоторые участки покровов тела затвердели, и в этих местах образовались внутренние выросты (так называемые подемы и апофизы), служащие для прикрепления мышц, участвующих в движениях частей тела и конечностей (см. также рис. 5.1, В, Г).
- Конечности нескольких посторальных сегментов приобрели функцию — хотя поначалу и не выраженную структурно) перемещения пищи к ротовому отверстию.
В результате всех этих преобразований возник характерный представитель типа Arthropoda.
Самыми ранними из известных членистоногих животных были трилобиты — обитатели морей, составляющие подтип Trilobita (рис. 2.5, 2.6). Они появились в кембрийский период примерно 600 млн. лет назад и исчезли примерно 200 млн. лет назад. У этих животных была небольшая голова, ориентированная вперёд, с антеннами и лабрумом, а позади головы несколько коротких ног, возможно участвовавших в захвате пищи. Ноги и их основания не были, однако, слиты с головой.
Трилобиты дали начало развитию нескольких морфологически различающихся групп. Возможно, что две из них послужили исходными для образования двух крупных групп членистоногих — подтипов Chelicerata и Mandibulata.
У Chelicerata антенны отсутствуют и тело состоит из двух частей — головогруди (цефалоторакса), выполняющей функции питания и локомоции, и брюшка (абдомена), выполняющего функцию размножения. К Chelicerata относятся скорпионы, пауки, клещи и близкие к ним группы.
У Mandibulata (ствол, ведущий к насекомым) первые три посторальных сегмента и их конечности слились с головой, которая состоит, таким образом, из первичного акрона и 6 сегментов тела. Конечности трёх сегментов, присоединившихся к голове, превратились в разнообразные структуры, предназначенные для захватывания пищи и помещения её в рот. Три сегмента, несущие эти ротовые придатки, называют гнатальными сегментами.
Такое слияние, объединившее в одну функциональную единицу все начальные структуры, связанные в конечном счёте с питанием, представляет собой характерную черту всех Mandibulata за очень редкими исключениями.
Древние Mandibulata дали начало двум линиям, каждая из которых представлена сегодня большой многообразной группой; это — Crustacea, или ракообразные (крабы, креветки и близкие к ним формы), и группа Myriapoda-Insecta (многоножки и насекомые). Ракообразные — преимущественно обитатели водной среды, как, возможно, и древние Mandibulata . Исходными же формами ветви Myriapoda-Insecta, вполне вероятно были наземные животные. Они могли жить в листовой подстилке по берегам мелких затенённых водоёмов и речек подобно некоторым современным Isopoda-Crustacea). Некоторые зоологи считают, что ракообразные стоят особняком среди современных Mandibulata, а сходвство в строении их ротового аппарата является результатом конвергентной эволюции.
В ветви Myriapoda-Insecta произошло слияние третьей пары ротовых конечностей, или придатков (которые у ракообразных называют вторыми максиллами), образовался длинный поперечный вырост — лабиум. Этим завершилось формирование новой и функционально важной стркутуры — предротовой, или преоральной полости, передняя и задняя стенки которой образованы лабрумом и лабиумом, а боковые — мандибулами и максиллами. Наличие ротовых придатков и предротовой полости обеспечивало животным большие возможности для захватывания пищи. Штёрмер (Stormer) оценивает это морфологическое приспособление как важнейшую адаптацию к наземной жизни.
Наземный предок многоножек и насекомых дал начало трём ветвям, сохранившимся и в наше время: одна представлена классом Chilopoda (губоногие), другая-тремя классами Symphyla, Pauropoda и Diplopoda (симфилы, пауроподы и двупарноногие), и третья — классом Insecta (насекомые). В первых четырёх классах (многоножки) ходильные ноги имеются почти на всех туловищных сегментах.
Насекомые подверглись дальнейшей специализации. Первые три пары локомоторных конечностей увеличились в размерах, остальные редуцировались, а затем исчезли совсем или превратились в специальные образования (рис. 2.1, Е), не связанные с локомоцией. Таким образом, локомоторная функция сосредоточилась в трёх первых сегментах, расположенных позади головы, и сформировался хорошо выраженный отдел тела, носящий название грудь (торакс). Задняя часть тела, где расположена большая часть органов, называется брюшком (абдоменом). Задние абдоминальные конечности превратились в наружные половые придатки, предназначенные для спаривания и откладки яиц. У некоторых ракообразных грудь и брюшко также чётко разделены, но при этом грудь обычно состоит примерно из 8 сегментов.
Анализ преобразований, имевших место в эволюции членистоногих от их примитивного безногого предка, позволяет предположить что а) неспециализированные конечности появились сначала на всех посторальных сегментах и б) затем подверглись постепенным изменениям, приведшим к их морфологической и функциональной дифференцировке. У насекомых это привело к чёткому разделению тела на три отдела: голову, несущую сенсорные придатки и ротовые части, грудь, несущую три пары ног, и брюшко, внутри которого находятся жизненно важные органы, а снаружи — терминальные придатки, выполняющие функцию наружных половых органов.
Представление о родственных связях внутри типа членистоногих, принятое в настоящем руководстве, схематически изображено на рис. 2.5. Согласно этому представлению, все гексаподы произошли от одной ветви древних членистоногих и поэтому представляют собой моно-филетическую группу. Как считает Кристенсен (Kristensen, 1975), предлагаемая схема происхождения насекомых нуждается в наименьшем числе эволюционных параллелей для своего объяснения. Другие авторы придерживаются иной точки зрения и считают, что не все насекомые произошли от одной эволюционной ветви, то есть их происхождение носит полифилетический характер (Manton, 1949, 1964, 1977). Обе гипотезы о происхождении насекомых рассматриваются в книге под редакцией Гупты (Gupta, 1979).
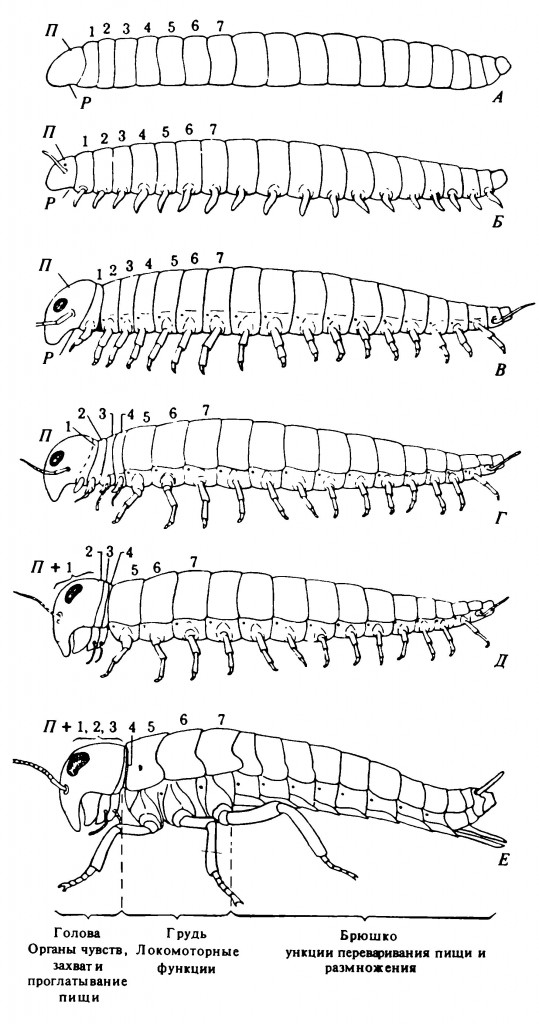
Рисунок 2.1. Схематическое изображение гипотетических стадий (А-Е) в развитии различных отделов тела и его придатков — от червеподобного предка до насекомого
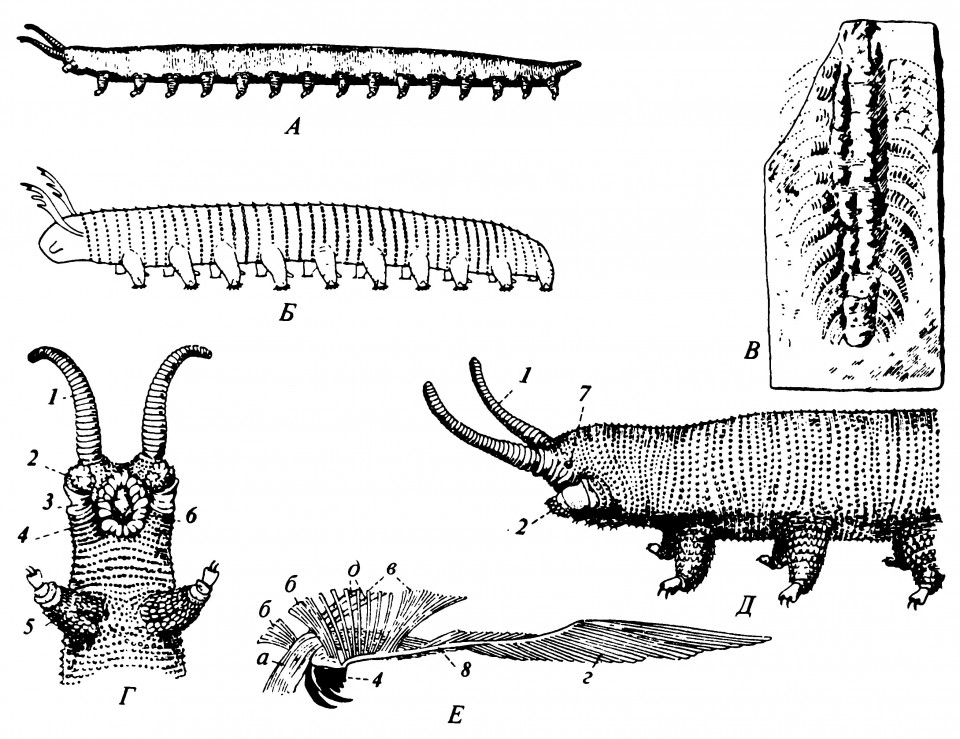
Рисунок 2.2. Древние и современные Onychophora
A. Peripatoides novae-zealandiae. Б. Реконструкция ископаемого Aysheaia pedunculata из среднего кембрия. В. Предполагаемый представитель онихофор Xenusion auerswaldae из докембрийского или раннекембрийского кварцита. Г, Д. Передний конец тела Peripatoides novae-zealandiae, вид с вентральной (Г) и латеральной (Д) сторон. Е. Его правая челюсть с мышцами: а-d — челюстные мышцы; 1 - антенна; 2 - оральная папилла; 3 - лабрум; 4 — челюсть; 5 - нога; 6 - складка вокруг ротового отверстия; 7 - глаз; 8 — аподема.
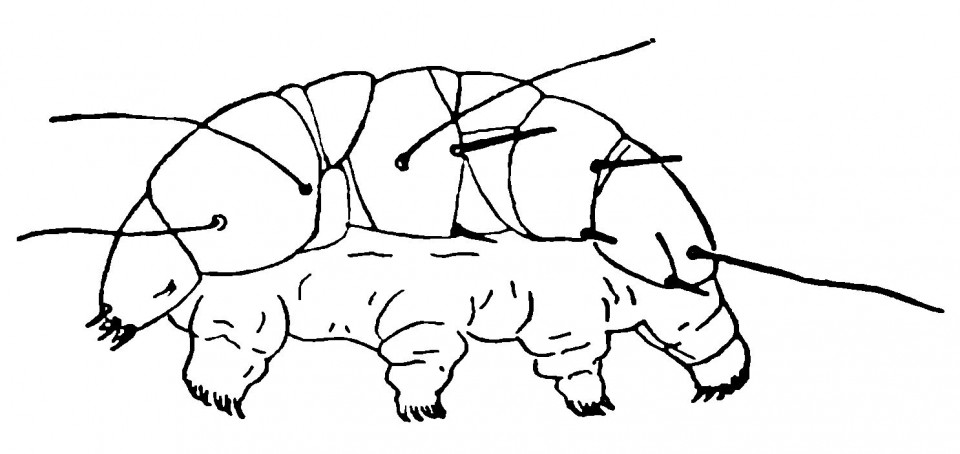
Рисунок 2.3. Тихоходка
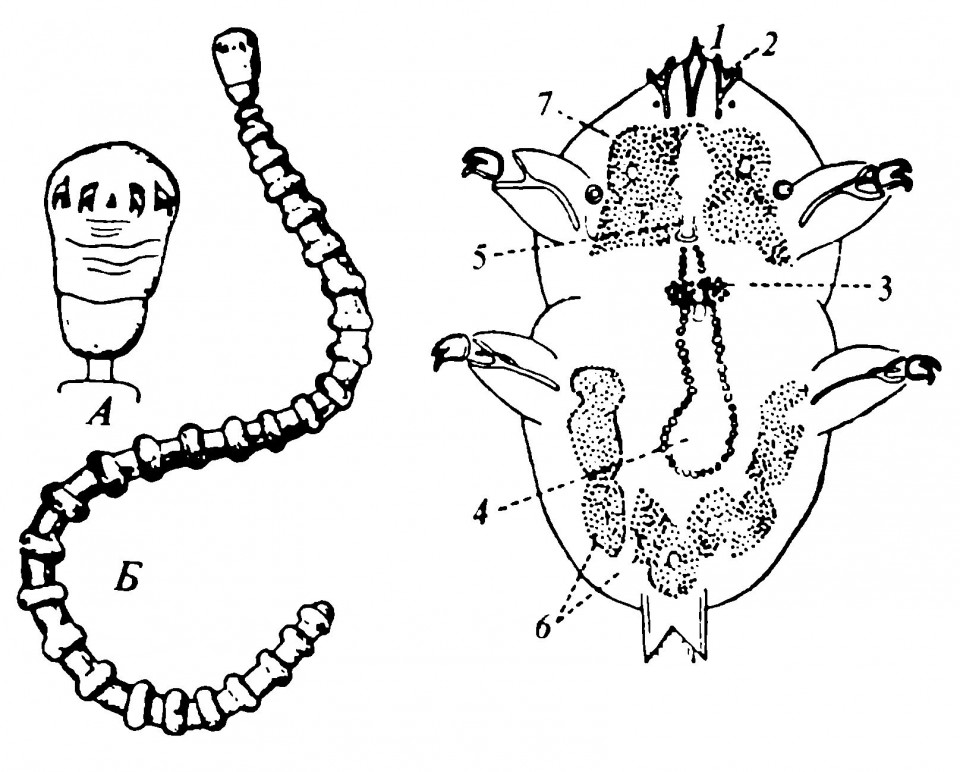
Рисунок 2.4. Пятиустка
Слева — Porocephalus annulatus. А. Голова, вид с вентральной стороны. Б. Целое животное, вид с вентральной стороны. Справа - личинка Porocephalus proboscideus, вид с вентральной стороны: 1 - вбуравливающийся передний конец; 2 - первая пара склеротизированных отростков, видимая между вилками второй пары; 3 - вентральный нервный ганглий; 4 - пищеварительный канал; 5 - рот; 6 и 7 - железистые клетки.
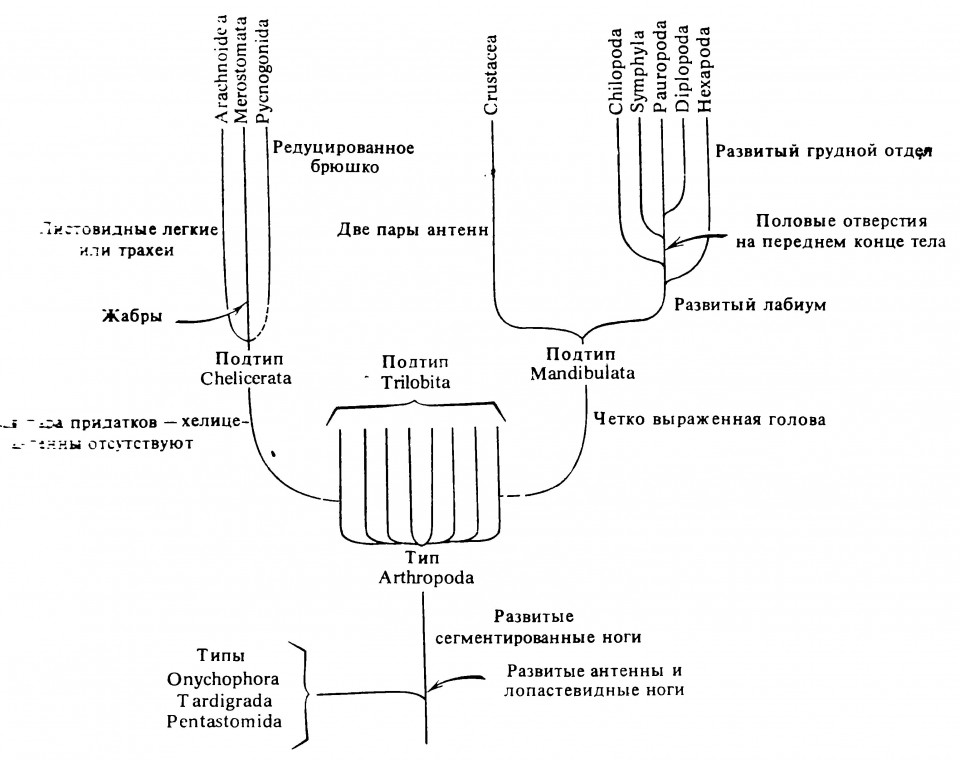
Рисунок 2.5. Гипотетическое родословное древо членистоногих
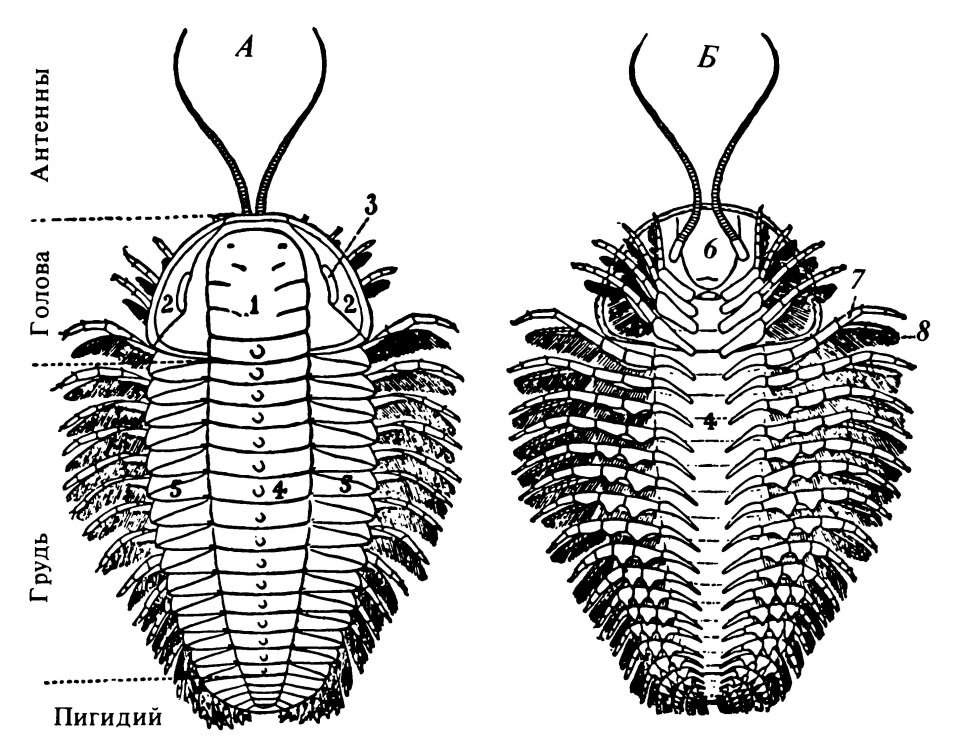
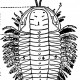
Рисунок 2.6. Общий вид трилобита Triarthrus becki
А. Дорсальная (верхняя) поверхность карапакса (щита), состоящего из трёх долей: плевры (5), осевой доли (рахиса, 4) глабеллы (1) и свободных щёк (2), несущих глаза (3). Б. Вентральная (нижняя) поверхность; видны двуветвистые конечности (7 и 8), прикреплённые к осевой доле, и верхняя губа, или гипостом (6), покрывающая ротовое отверстие. Двуветвиртые ноги имеют двойную функцию; верхняя опушённая ветвь несёт жабры для дыхания и служит, кроме того, гребными лопастями, нижняя голая ветвь служит для ползания. Короткие передние придатки, возможно, участвуют в процессе питания.

Arthropoda: насекомые и близкие к ним группы