Alarm Pheromones of Insects
Alarm pheromones are defined as chemical substances, produced and released by an organism, that warn or alert another of the same species of impending danger. This is exemplified by many species of aphids (Hemiptera: Aphididae) in which the pheromone is caused to be released by attack, for example, by predators, with ensuing dispersal by which individual aphids may avoid a subsequent attack. However, the term alarm pheromone also is employed when the responding individuals are stimulated to show aggression towards the attacking agent. This is common in the social Hymenoptera; for example, the honeybee, Apis mellifera (Hymenoptera: Apidae), and many ant species respond aggressively to their alarm pheromones.
As alarm pheromones can benefit the survival of members of the species involved, it is common for insects that employ alarm pheromones to live in congregations for some or all of their life cycle. In the case of social Hymenoptera, the colony is genetically related, and in asexually reproducing aphids, the colony is clonal. Although the survival of siblings or clones by alarm pheromone response at the cost of the attacked individual appears altruistic, in genetically related colonies, genes from the individual will predominate in the survivors and be passed on to their kin.
Hemipteran Alarm Pheromones
Alarm Pheromones of Aphids (Hemiptera: Aphididae)
When disturbed or attacked, many aphid species release alarm pheromone from droplets secreted from tube-like structures called cornicles on their dorsal posterior. This phenomenon has been studied exclusively in the asexual forms and most often in asexually reproducing wingless females. Aphids nearby exhibit a variety of behaviors ranging from stopping feeding and moving away, to running or dropping off the plant and even attacking the predator. However, not all aphids in a group respond. The relative risks of predation and costs of escape, for example, cessation of feeding and risk of desiccation, affect the likelihood of any particular response. In studies of the peach-potato aphid, Myzus persicae, the pea aphid, Acyrthosiphon pisum, and the rose-grain aphid, Metopolophium dirhodum, early stages were found to be less sensitive to alarm pheromone than later ones.
However, older wingless M. persicae require the greatest stimulation of alarm pheromone before responding, while winged M. persicae, particularly those not feeding, are extremely sensitive to alarm pheromone. The lack of response from the early stages suggests that the risk of predation to these nymphs is lower than the risk involved in ceasing to feed and dropping from the plant. When young M. dirhodum respond to alarm pheromone, they do so by moving to another part of the plant rather than by dropping. Winged adults, on the other hand, are more responsive to alarm pheromone, perhaps because they can more readily move off the host. The sugarcane woolly aphid, Ceratovacuna lanigera, also shows different reactions to alarm pheromones at different life stages; it shows attack behavior until adult, when the normal aphid dispersal response takes over.
Considerable variation is seen between aphid species in their sensitivity to alarm pheromones and in both the speed and the form of the response. This variation often can be explained by differences in the ecology of the species. Some aphids, particularly those tended by ants, stay on the plant and respond by walking or “waggling” their abdomens rather than falling off the plant. These aphids appear to depend more on the protection afforded by their ant attendants than their own defensive mechanisms. Aphids that walk away from a source of alarm pheromone tend to form new clusters a short distance from the original site, thus ensuring continued ant attendance.
Susceptibility to insecticide also has been found to correlate with responses to alarm pheromone. Susceptible strains produce more pheromone and respond more quickly and in higher numbers than insecticide-resistant strains. In addition, clones collected from around the world showing knockdown resistance to pyrethroid insecticides, and esterasebased insecticide resistance, showed lower levels of disturbance to the synthetic alarm pheromone. These aphids may therefore suffer increased predation or parasitism in the absence of insecticides, affecting the evolutionary fitness of insecticide resistant clones. This may be due to physiological effects associated with resistance, which could affect mobility or sensitivity of the nervous system to stimuli.
Aphids that have dropped from a plant may re-colonize or may move to another host plant further away. The turnip aphid, Lipaphis erysimi, and M. persicae dislodged by alarm pheromone are less likely to return to the original host plant than when mechanically dislodged. Similar patterns of behavior are found in A. pisum. Aphids dislodged by a predator or experimentally with synthetic alarm pheromone spend longer “running” before the “search” for a host plant began, whereas aphids dislodged mechanically are more likely to begin to search for a host plant immediately.
Droplets secreted from the cornicles comprise two types of material: a volatile, rapidly vaporizing fraction which is the alarm pheromone, and a waxy fraction, consisting mainly of triglycerides, that crystallizes on contact with foreign particles outside of the aphid’s body. The waxy component appears to function as a sticky or quick-setting irritant to predators and parasitoids and a releasing substrate for the alarm pheromone component.
The main component of the alarm pheromone (Fig. 30) of many aphids is the sesquiterpene hydrocarbon (E)-β -farnesene (1). Other components may also be present as found in the alarm pheromone blend of the vetch aphid, Megoura viciae, which contains the monoterpenes (-)-α pinene, (-)-β -pinene, (Z,E)-α -farnesene and (E,E)-α -farnesene, in addition to (E)-β -farnesene. There is a high degree of cross-activity of both natural alarm pheromone and (E)-β -farnesene among species within the aphid subfamilies, Aphidinae and Chaitophorinae. This is typical of insect alarm pheromones in general, since such cross-activity does not reduce their evolutionary value. However, the main component of the alarm pheromone of the spotted alfalfa aphid, Therioaphis maculata, and the sweet clover aphid, T. riehmi, in the Drepanosiphinae, is the cyclic sesquiterpene (-)-germacrene-A (2).
In the turnip aphid, L. erysimi, it has been demonstrated that isothiocyanates, acquired from chemicals in the host plants, synergize the effect of the alarm pheromone. These isothiocyanates are likely to be released from aphid honeydew so that, when there is a high number of other aphids in the immediate vicinity, the percentage of aphids responding to alarm pheromone increases.
Alarm Pheromones of True Bugs (Hemiptera, Heteroptera)
The Pentatomidae is the dominant family of stinkbugs, or shield bugs. The family comprises many species that are pests of economic importance, especially in warmer climates. These insects secrete a complex mixture of chemicals when strongly molested. The energetic cost of the defense response, especially production of defense chemicals, is significant and considerable provocation usually is required to cause release. In adults, the source of the defense compounds is the metathoracic gland, while in nymphs it is the dorsal abdominal glands. These are precursors to the metathoracic gland in adults and perform the same defense function. The chemical content of these secretions is similar throughout the order; for example, the components of the secretion of the stinkbug Cosmopepla bimaculata are a complex mixture of hydrocarbons, aldehydes and esters. The secretion, which can be ejected from either or both metathoracic glands in controlled amounts or even resorbed, displays a defensive function as a predator repellent. In one case, researchers have shown uncommon dedication in describing the repellency by squeezing adults in their mouths and chewing nymphs. The effects were a burning sensation and numbness of the tongue for up to two hours. In addition to repelling predators, the secretions possess alarm pheromone activity and cause adults to drop off plants. In the field, C. bimaculata are found highly clumped and the occurrence of large numbers living together gives an evolutionary advantage to possessing an alarm pheromone.
Six-carbon-long aldehydes, in particular (E)2-hexenal, are common components of defensive secretions and are found in many families of heteropterous insects, including the Pentatomidae, Coreidae, Pyrrhocoridae, Cimicidae, Cynidae and Alydidae. it is thought that the general irritant properties of aldehydes provide a repellent effect to predators with hydrocarbons such as n-tridecane, another ubiquitous component, acting to spread the oily secretion so that the aldehydic components can exert full irritant effect. (E)-2Hexenal has been reported to have the added dual functions of both alarm and aggregation pheromone, depending on the stimulus concentration, as well as use as a defense chemical. In the case of the bed bug, Cimex lectularius, and Eurydema rugosa, low concentration of (E)-2-hexenal acts as an aggregation pheromone while high concentration produces an alarm response. Alternatively, it has been reported that n-tridecane, the other ubiquitous component of defense secretions, is a bifunctional pheromone for the southern green stinkbug, Nezara viridula (Heteroptera: Pentatomidae), which causes dispersal at high concentration (one individual equivalent) and aggregation at low concentrations. The multifunctional aspect of these compounds has important repercussions for their practical use as dispersal agents for pest species in the field, as the low concentration response of aggregation may dominate once the applied high concentration of compounds has diminished.
The pentatomid bug Erthesina fullo is a major pest of pine and hardwood trees. Both sexes produce a secretion from the metathoracic gland causing conspecific adults to drop from plants, fly or move away. The secretion comprises nine identified compounds, esters and aldehydes (about 35%) including (E)-2-hexenal and (E)-4oxo-2-hexenal (3) and long chain alkanes, including 50% n-tridecane. Likewise, adult and nymph secretions from Dysdercus cingulatus (Heteroptera: Pyrrhocoridae) revealed 55 identified compounds, although the major components are again aldehydes and n-tridecane, features common with several more species of pentatomids from the genus Chlorochroa and Piezodorus guildinii.
The leaf-footed bug, Leptoglossus zonatus (Heteroptera: Coreidae), is an economically important pest of Brazilian corn. An extract obtained from the metathoracic gland by immersion in hexane showed that the major compounds were all of six-carbon length: hexanal, hexanol, hexyl acetate, hexanoic acid, and (E)-4oxo-2-hexenal (3). (E)-2-Hexenal was found in the nymph extracts but not in the adult, an example of the general rule that exocrine chemistry of heteropterous nymphs is distinct from that of the adult. In this case, different life-stages possess different alarm pheromone systems. When tested individually, all components produced varying degrees of alarm response in adults and nymphs and even mating insects would stop and disperse, over-riding the sex pheromone response. These compounds are not species specific and are, for instance, found in L. oppositus and L. clypealis, a situation that mirrors the cross-activity of (E)-β -farnesene in many aphid species (see above), providing more evidence that this non-specific activity does not reduce alarm pheromone value in evolutionary terms.
Adults of the bean bug, Riptortus clavatus (Heteroptera: Alydidae), a pest of Japanese soybean, secrete (E)-2-hexenyl acetate in its defensive response. This causes an alarm response in adults and nymphs. Interestingly, adults also produce (E)-2-hexenal, and although some response was found when tested at high concentration, there was no response at physiological concentration, suggesting that this compound is not an alarm pheromone.
The examples shown above demonstrate that alarm pheromones of heteropteran families are based on a chemical selection general to insects of a wide taxonomy and show little species specificity. The alarm behavior caused by high concentrations of n-tridecane or (E)-2hexenal is rationalized easily but the aggregation due to low concentrations is more difficult to explain. Perhaps these substances are constantly emitted in very small quantities due to their volatility and act so as to direct individuals to a region where conspecifics can be found and therefore where food most probably is located, as well as the defensive advantage in being part of a large group.
Triatomine bugs (Heteroptera: Reduviidae) are blood-sucking insects that live throughout the Americas and cause public health problems by transmitting the protozoa Trypanosoma cruzi, the causative agent of Chagas disease, to humans. Secretions from Brindley’s gland (a simple sac, metathoracic in origin) of several species all revealed isobutyric acid as the major component. Subsequently, other short-chain and branchedchain fatty acids have been identified, and together with isobutyric acid, they act as a powerful defensive secretion. Pure isobutyric acid vapor, however, also caused an alarm response in Rhodnius prolixus while another report revealed that low concentrations of isobutyric acid attracted R. prolixus adults. This defense compound therefore shows the same multifunctional alarm and aggregation properties as described for components of the stinkbug secretions (see above). Triatomine bugs are inactive and hide during the day, congregating in protective sites. This aspect of group living can help explain the evolutionary advantage in possessing aggregation and alarm responses.
Alarm Pheromones of Social Insects (Hymenoptera)
The Honeybee (Hymenoptera: Apidae)
When the honeybee (Apis mellifera) is attacked, alarm pheromones released serve to muster help and to direct the attack. Specialized guard bees present at the nest entrance carry out attacks. Although these guards are relatively few compared to the colony population, release of alarm pheromone can result in synchronized attacks by more than 100 workers against an intruder. Guard bees initiate attacks by raising their abdomens, protruding their stings and releasing alarm pheromone from the sting chamber. The workers then alert the rest of the hive by wing beating, aiding dispersal of the pheromone, and by running into the hive. Aſter a few seconds, many excited bees may rush out of the hive entrance and search, or stop and assume a characteristic tense and aggressive posture with a slightly raised body, wings extended, mandibles agape and antennae waving. In this highly activated state, they will fly to attack at the slightest further provocation. These two stages of alarm response are called alerting and activation and are characteristic of alarm pheromones.
Alerted workers need to search for and discover the enemy to prevent any further threat. To do this, they rely on other cues to direct the attack such as odor, jerky movement and hairy body covering. Once the threat is located, it is stung, injecting a dose of venom. However, the shaft of a sting is barbed and a bee is unable to withdraw it from the skin of vertebrates, so the sting, together with associated motor apparatus and glands, are severed from the bee as it attempts to fly away and are leſt attached to the enemy. The severed sting apparatus continues to pump venom into the victim and alarm pheromone is dispersed from the exposed under-surface of the sting shaft membrane to mark an enemy and make it a more obvious target.
The main alarm pheromone component of the sting gland was identified in 1962 as isopentyl acetate (4). Although a number of other compounds are known to be present, isopentyl acetate and (Z)-11-eicosen-1-ol (5) account fully for the activity of the sting pheromone. The roles of the two compounds in the pheromone appear to differ, with (Z)-11-eicosen-1-ol responsible for prolonging the activity of isopentyl acetate. Other compounds such as
Stinging bees often grip an enemy with their mandibles and deposit an alarm substance. At the hive entrance, more bees examine mandibular gland extracts of worker honeybees applied to filter paper than examined unscented filter paper.
Under certain circumstances, honeybee alarm pheromones are repellent. The presence of alarm pheromone deters honeybees from foraging at dishes of sugar syrup and from exposing their Nasonov glands and fanning which normally attracts other bees. Furthermore, a high concentration of alarm pheromone repelled foraging bees from crops including oilseed rape, normally highly attractive to bees, in an area that had many honeybee colonies.
Alarm Pheromones of Ants (Hymenoptera: Formicidae)
The Formicidae is a huge family comprising thousands of ant species, all of which are social insects, living in colonies that vary hugely in size. Members of an ant colony may be differentiated into castes that specialize in carrying out particular tasks and vary in their response to alarm pheromone. Soldiers show a more aggressive response, are more likely to respond when the threat is closer to the nest and may be specialized to deal with vertebrate predators. Also, workers of the Texas leaf-cutting ant, Atta texana (Formicidae: Myrmicinae), have a lower threshold for alarm pheromone response than the queen and males. Other factors governing the type and intensity of alarm response are the age and size of ant colony. When alarm pheromone is present in sufficient concentration to excite the workers, other stimuli are needed to direct an attack. Workers often will touch everything they encounter and the full-scale alarm response may rely on additional cues, such as the presence of an alien object. Alarm pheromones also may function with acoustic alarm signals. Ant species in the sub-family Dolichoderinae produce vibration signals using their mandibles to scratch the ground or the abdomen to hit the ground, increasing alarm behavior in other workers. Vibrations are produced also by leaf-cutting ants which act as warning signals. Alarm pheromones also are used by ants to attract attention if they are trapped, and may be released by reproductive ants just before mating flights to ensure that aggressive workers protect them from potential predators.
The context in which a worker encounters an alarm pheromone also influences the response. Workers of the grass-cutting ant Atta capiguara (Formicidae: Myrmicinae) are less likely to show alarm behavior if already engaged in a task. Foragers carrying leaves do not respond to alarm pheromone, whereas minor workers and foragers that are not carrying leaves do respond.
Ant alarm pheromones may be produced from one or several sources. The army ants or Eciton ants (Formicidae: Ecitoninae) and the rare Leptanilla sp. (Formicidae: Leptanillinae) of Indonesia have large mandibular glands, which are believed to be the sole source of the alarm pheromone. However, other species rely on a combination of secretions from several glands. Formica and Myrmica species use products from the poison and Dufour’s gland (both opening near the base of the sting), as well as the mandibular gland. Ponerine ants (Formicidae: Ponerinae) use secretions from the pygidial gland as alarm components, whereas the poison gland is the most important gland of several other species, including the harvester ant, Messor barbarus (Formicidae: Myrmicinae).
Ants are able to detect and respond to specific isomers of their alarm pheromone. Myrmica rubra and M. scabrinodis use
Myrmica species of grass-cutting ants share the main component of alarm pheromone, 4- methyl-3-heptanone (9), but they have speciesspecific modifying components. The response to 4-methyl-3- heptanone was compared to that elicited by the bodies of workers that had their heads crushed to release the natural alarm pheromone. 4-Methyl-3-heptanone and bodies caused the same level of attraction but the full range of alarm behavior was seen only with the bodies. In contrast, workers of the giant tropical ant, Paraponara clavata (Formicidae: Paraponera), produce two components, 4-methyl-3-heptanone and 4-methyl3-heptanol.
Atta capiguara is a grass-cutting ant species that lives in colonies with hundreds of thousands of workers. Workers are polymorphic, varying in size from small minors and medias to the larger foragers and soldiers. Minors and medias do most of the nest tasks whereas foragers collect the grass; however, minors often are found on foraging trails despite the fact they do not carry grass. They are believed to be patrollers as they have a stronger response to alarm pheromone than foragers and soldiers. Minors of other Atta species are also more efficient at recognizing intruding ants than other castes.
As is the case with honeybees described above, the complete alarm response can be described by a number of behaviors. These behaviors and their elicitors have been dissected in an elegant piece of research on the African weaver ant, Oecophylla longinoda. The major workers produce a secretion from the mandibular gland comprising four active components: hexanal, hexanol, (E)-2-butyl-2octenal (10), a dimer of hexanal produced chemically by self-condensation, and
The properties that make alarm pheromone cues for conspecifics also enable them to act as cues for parasites and predators of ants to find their prey. Apocephalus paraponerae is a parasitic fly that attacks the ant Paraponera clavata. Females and males of A. paraponerae are attracted to injured, fighting or freshly killed workers. Aſter finding a worker, the female lays a few eggs, which will hatch and then feed on the victim for
Alarm Pheromones of Thrips (Thysanoptera: Thripidae)
Thrips are small, economically important pest insects, often known as thunderflies. The defensive behavior of thrips includes raising and lowering the abdomen and secretion of a droplet of anal fluid highly repellent to predatory ants. Western flower thrips, Frankliniella occidentalis (Thysanoptera: Thripidae), are not social but tend to be found in clumped distributions. Adults and nymphs of western flower thrips produce an anal droplet containing decyl acetate and dodecyl acetate (13) in a molar ratio of 1.5:1. Each component, at levels of 1 ng, produces the alarm response of walking away from the source or dropping from leaves. The response, however, is only over short distances and limits the potential for pheromone use in pest management.
Cockroach Alarm Pheromones (Blattodea: Blattidae)
Defensive secretions are well known in cockroaches. They are produced from ventral inter-segmental glands and comprise an organic and an aqueous phase. In the case of the Florida woods cockroach, Eurycotis floridana (Dictyoptera: Blattidae), 90% of the organic phase (which comprises 85% of the total secretion) is (E)-2-hexenal, a compound found in many heteropteran bugs and discussed above. The rest of the organic secretion comprises approximately 40 other components, including mainly aldehydes, alcohols and carboxylic acids, while the aqueous phase contains gluconic acid, glucose and gluconolactone. The secretion acts as a conspecific alarm pheromone in these gregarious insects with nymphs responding at lower concentrations than adults do. Ethanolic extracts of the American cockroach, Periplaneta americana, also repel conspecifics from aggregations in daytime shelters. However, there was no evidence that this repellent is released by living insects as an alarm pheromone but is instead endogenously produced from dead insects and is effective against other cockroaches with diverse phylogenetic relationships. The effect, therefore, is not pheromonal, as the authors explain the activity in terms of unsaturated fatty acids (oleic, linoleic and linolenic acids) which emerge as signals of death and injury among organisms from a wide phylogenetic background. Both of the reports described above provide evidence that the use of alarm pheromones to increase dispersal for pest management purposes will be of limited value. Due to the aggregation effect at low concentration, treated areas could become attractive. Also, if low concentrations were used as attractants in a lure and kill approach, dead insects would repel others before they become ensnared.
Alarm Pheromones of Beetles (Coleoptera)
Despite the vast numbers of species in the order Coleoptera, inhabiting a wide range of ecological niches, little is known of the existence of behavioral responses to alarm pheromones they may possess. Species of beetles that are group-living are most likely to demonstrate alarm responses. Gyrinid beetles (Coleoptera: Gyrinidae), known as whirligig beetles, live in open habitats on fresh water surfaces and typically aggregate in groups containing hundreds of individuals, dispersing in the evening to forage. Although easy to detect by fish, they are seldom preyed upon due to a repellent secretion released as a last resort to physical attack. The secretion also acts as an alarm pheromone over short distances, increasing locomotory activity and defensive movement, such as diving and active underwater swimming. Although alarm dispersal aſter attack can occur, the aggregations of beetles themselves indicate to experienced predators to avoid the group and confer an aposematic effect acting at the group level rather than the individual level.
Lacewing Alarm Pheromones (Neuroptera: Chrysopidae)
The green lacewing, Chrysoperla carnea (Neuroptera: Chrysopidae), is an important predator of pest aphids and, as such, is a beneficial insect. It discharges a malodorous secretion from glands at the anterior of the prothorax. The major component of this secretion has been identified by gas chromatography, mass spectroscopy and chemical synthesis as (Z)-4-tridecene (14) and gas chromatography coupled electroantennagrams revealed that it is detected by the lacewing antennae. Predatory ants displayed avoidance behavior in response to it, suggesting a defensive function, and in laboratory experiments, adult lacewings avoid entering areas where it is present. In the field, it acts as an antagonist to trap catches using known attractants and, as such, could be described as an alarm pheromone. Another species of lacewing, Peyerimhoffina gracilis, also produces the identical compound. As lacewings are not known to be gregarious, the exact ecological purpose of this compound is being investigated.
Conclusions
It can be seen that alarm pheromones are used widely by a broad taxonomic diversity of insects and elicit equally varied behavioral responses, including escape or aggressive behavior. Alarm pheromones are generally low molecular weight, organic compounds and so are volatile, dispersing quickly, and do not persist in the environment. In addition, the chemical nature of the alarm pheromone often is unstable, increasing the lack of persistence. This allows conspecifics to be alerted very quickly over a fairly large area and yet not cause false alarm aſter the danger has passed. Alarm pheromones are often produced in glands responsible for biosynthesis, storage or release of defense secretions. This association between alarm pheromones and defense glands, including those near the sting or mandibles, has led to the hypothesis that alarm pheromones have evolved from chemicals that originally had a defensive role, or are themselves defense compounds that have taken on an additional alarm pheromonal role. The fact that known defense components have additional multifunctional pheromonal roles of alarm (high concentration) and aggregation (low concentration) also points to the possibility that these pheromonal roles have evolved from compounds originally used for defense. Of particular interest is the common lack of species specificity found in alarm pheromones, which is in contrast to that of other pheromones.
Sex pheromones, for example, are so specific that they can be the sole identifiable trait in defining morphologically identical populations, such as within the species complex of the sandfly, Lutzomyia longipalpis (Diptera: Psychodidae). However, the alarm pheromone of different aphid species is (E)-β -farnesene and different species of Atta grasscutting ant use 4-methyl-3-heptanone. In addition, production of (E)-2-hexenal and n-tridecane is ubiquitous as multifunctional pheromone components in terrestrial true bugs, and it is possible that (E)-4-tridecene may reveal itself to be common in green lacewings. Discrimina tion between these behavioral signal compounds, therefore, is not essential to their function as alarm pheromones, and there may even be evolutionary benefits in being able to respond to alarm pheromones of related species of insects.
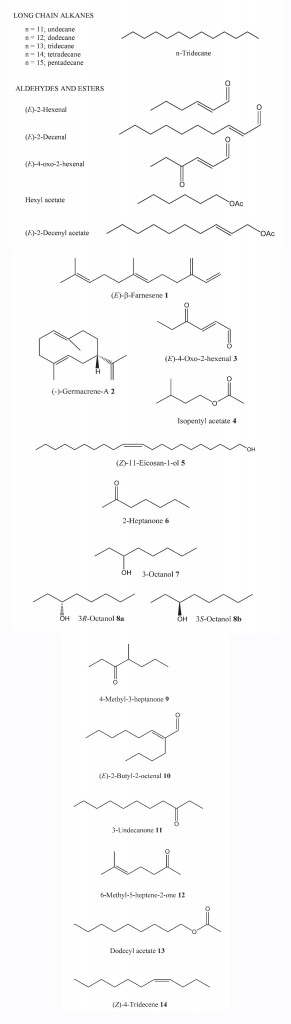
Figure 30 Defense secretion of Cosmopepla bimaculata showing a typical range of compounds produced by stinkbugs

Agroecology