Acoustic Communication in Insects
Sound is used by a wide variety of insects for diverse purposes. It is difficult to evolve an acoustic communication system. There must be modifications to produce sound, transmit the sound in the environment and modify the sound to specific biological purposes, as well as the evolution of structures that will receive and decipher the signals. Arthropods are one of only two major groups of animals (along with the vertebrates) that have evolved acoustic communication and insects are the primary group of arthropods to exhibit acoustic behavior. At least ten orders of insects possess species that produce acoustic signals.
Sound
Sound is generated by causing the repeated compression and rarefaction of an elastic medium. The waves of compression produced will then travel through the medium to the receptor or target. The medium can be air, water or a substrate — so sound also encompasses vibrations. As the sound energy travels through the environment, it is modified by various interactions with the components of that environment. In addition, sound waves are reflected as they move through any environment. This bending of sound waves can initiate interactions among the waves. The signal begins to deteriorate as a result of these wave interactions and with the variations in the signal initiated due to temperature and humidity. Acoustic energy is also lost as it is absorbed by structures in the environment and as a result of spherical spreading from the sound source. The loss of signal integrity due to these environmental interactions is termed the excess attenuation of the signal. The amount of excess attenuation varies with the habitat and the original signal properties.
To complicate matters, the small size of insects requires that they use high frequencies (Fig. 18) to transfer energy efficiently to the signal. This will limit the range of a given signal because higher frequencies attenuate more rapidly in the environment. To use lower frequencies, insects must modify their bodies or behaviors to increase the efficiency of sound production. For example, tree crickets (Gryllidae) create a baffle by inserting their body in a leaf to decrease the acoustic short circuiting of a small dipole sound source, mole crickets (Gryllotalpidae) dig burrows which act as loudspeakers that amplify and direct their calls skyward, and the hollow abdomen of male cicadas (Cicadidae) acts as a resonating structure to amplify the acoustic signals produced by the tymbals.
The sound producing systems of insects are generally vibrating structures. These structures are necessary because muscles cannot contract rapidly enough to produce high frequency sounds. The sound generating structures vibrate multiple times for each muscle contraction, so the sound producing systems act as frequency multipliers. Once the vibrations are initiated, the sound can be modified by attached resonant structures.
There are many different schemes that can be employed to describe insect calls. The variety of sound production mechanisms has led to a variety of terminologies. No one terminology has been successful in describing the different sound production mechanisms or phylogenetic relationships of insects. When sounds are recorded and analyzed in any acoustic work, each analysis of acoustic signals in
Sound Production
There are several different types of sound production mechanisms that have evolved in insects. The relatively low muscle contraction frequency means that additional structures had to evolve in order to produce the high frequency sounds that small body size will require to generate acoustic signals efficiently.
The primary mechanism used by insects to produce sound is a stridulatory apparatus. The chitinous exoskeleton and jointed appendages of insects are preadapted for modification into stridulatory apparati. Each stridulatory apparatus is composed of a file and plectrum or scraper. The file is generally a row of small cuticular teeth that is rubbed against a ridge or blade (the plectrum) on some other body part (Fig. 19). The teeth are bent as they catch on the plectrum and pop forward as they release from the plectrum. The release causes the teeth to vibrate alternately compressing and expanding air molecules producing sound. In general, these vibrations will occur in a single plane, resulting in a dipole being formed that produces an asymmetrical sound field at close range. The joints and exoskeleton of insects have permitted stridulatory apparati to evolve in many locations on the body.
There are file and scrapers found between antennal segments (Phasmatodea), separate mouthparts (Orthoptera), head and thorax (Coleoptera), abdominal segments (Hymenoptera), wings and thorax (Lepidoptera, Hemiptera), body parts and legs (Hemiptera, Orthoptera), legs and wings (Orthoptera, Lepidoptera, Coleoptera), legs and legs (Thysanoptera, Hemiptera), between wings (Orthoptera), and between segments on the genitalia (Lepidoptera, Hemiptera). A tymbal organ is a specialized sound production organ. It is a ribbed, chitinous membrane attached to a tymbal muscle (Fig. 20). The tymbal buckles as the tymbal muscle contracts. Sound pulses are produced when the tymbal buckles, when individual ribs buckle and potentially when the tymbal returns to its relaxed position. The unbuckling of the tymbal is assisted by the resilin within the tymbal. The resonant frequency of the tymbal determines the frequency of the sound pulses produced. Additional structures such as the opercula, tymbal covers, various muscles and the abdominal air sacs can modify the sound produced. The abdominal air sacs can become rather large as in the bladder cicada Cystosoma saundersii Westwood whose abdomen is so large, to resonate at a low frequency, that the male has difficulty flying. The tymbal is a common organ in the Hemiptera and acts as the ultrasonic pulse generator in the Lepidoptera.
Percussion is another mechanism of sound production used by insects. Crepitation, a clicking sound produced by the wings, is another percussion mechanism in insects. The wings are clapped together or banged on the substrate to produce a sound pulse in some Hemiptera. It is relatively rare for airborne signals which are generally produced as a by-product of flight. The wings may strike each other or the legs during flight-producing sound pulses as in the acridid grasshoppers (Orthoptera). However, specialized percussive sound production systems have evolved. A castanet is found on the costal margin of the tegmina in some moths (Lepidoptera) which produces a pure tone when struck. The acridid grasshopper Paratylotropidia brunneri Scudder can snap their mandibles together to generate sound. Percussion has been observed in Hemiptera, Isoptera, Plecoptera, Lepidoptera, and Orthoptera.
Tremulation is the vibration of unspecialized body parts to generate sound. The abdomen is oſten moved dorso-ventrally or laterally to send vibrations through the legs to the substrate. This mechanism has been well studied in the lacewings (Neuroptera). Wing vibrations can also be used to send information over short distances. This type of signal is produced by members of the Diptera (flies and mosquitoes) and Hymenoptera.
The expulsion of tracheal air is the final and relatively rare mechanism of insect sound production in insects. This method of sound production has been described in a number of insects, most notably the Madagascan hiss ing cockroach (Gromphadorhina portentosa [Schaum]), the Death’s Head hawk moth (Acherontia atropos [L.]) and some African Sphingids (Lepidoptera).
In addition to the airborne signals produced by acoustic insects, it has now been shown that vibrational signals are also produced by sending information through the substrate. These vibrational signals are primarily produced by tremulation but also can be important in deciphering an airborne signal, particularly at close range to the sound source. Vibrational signals have been identified in members of the Neuroptera, Diptera, Hemiptera, Plecoptera, Coleoptera, Orthoptera, Mecoptera, Raphidioptera, Lepidoptera and Hymenoptera.
Sound Reception
A receptor for acoustic signals is necessary for a communication channel to exist. Because sound is produced by changing air pressure, a modified mechanoreceptor is needed to sense the acoustic information. The receptors can be classified as either a pressure detector or a particle detector. Pressure detectors are membranes that bend when pressure is unequal on the two sides of the detector. Particle detectors are long structures that move when impacted by many particles moving in the same direction. The movement of the particle detector alternately stretches and compresses sensory cells at the base of the organ.
The primary type of pressure receptor organ is a tympanum, which has evolved independently in at least eight orders of insects. Tympana are generally a thin membrane system stretched over a closed cavity. The tympanum bends away from the side of higher pressure setting up oscillations as the sound waves strike the membrane. Each tympanum has a resonant frequency based on its thickness, size and shape. The resonant frequency is generally tuned to the carrier frequencies of the communication channel to increase the efficiency of information transfer.
The tympana are associated with other mechanoreceptor organs to transduce the signal for the nervous system. As the tympana oscillate, vibrations are sent to various types of receptors, generally a specialized chordotonal organ called a scolopidial organ, which act as the input site to the central nervous system. There can be elaborate structures associated with the tympana to transduce energy into the central nervous systems such as the crista acoustica of the crickets and katydids (Orthoptera) and Müller’s organ of locusts (Orthoptera), which provides frequency discriminating abilities. These pressure receptors are generally found in pairs, one on each side of the body. This provides an animal with a means to determine the direction from which the sound originated.
The sensory structures for vibrational signals are trichoid sensilla generally found on the feet or cerci or through specialized subgenual and metatarsal organs located in the legs. The subgenual organs are located just distal to the femoral-tibial joint in all six legs to promote detection and directional hearing. They are similar to tympana in that they are chordotonal organs but lack the tympanal membranes and tracheal air sacs of the tympanal system.
Particle receptors are generally specialized structures found in specific groups of insects. For example, female fruit flies (Diptera) sense the courtship signals produced by the male with a special antennal segment called an arista. Male mosquitoes and midges (Diptera) have plumose antennae which detect the species-specific wing beat frequencies produced by females. Johnston’s organ is a more complex chordotonal organ found in the antennal pedicel of some Diptera and Hymenoptera which is stimulated by the vibration of the antennal flagella. The long bristles and antennae provide a mechanical advantage to the sensory cells increasing the sensitivity of the receptor.
Functions of Acoustic Signals
Sound has many potential benefits in that the signals can be used day or night, can be modified quickly, and can travel a significant distance even if conditions are not optimal for other signal pathways. However, sound is energetically costly to produce and advertises your position to potential predators as well as potential mates. Even with these potential problems, insects have evolved diverse functions for acoustic signals. Acoustic signals are used for a variety of purposes in insects including sexual signaling, courtship signals, aggression, social recruitment and defense.
A primary function of acoustic signals is as an intraspecific communication channel. Sounds are used to attract mates and to isolate species reproductively. Each species has a characteristic call that can prevent related species from cross mating. The calls produced by individual signalers can be compared by receivers providing the opportunity to select a mate who is producing a call that exhibits specific characteristics. This is particularly true when callers have congregated into localized areas, which is another function of acoustic signaling. Mates may be selected based on the number of calls, the temporal patterns, loudness, etc. The specific characteristics used by a given species are usually chosen based on their ability to demonstrate the viability of the caller. Duets between individuals (either intrasexual or intersexual) can also act as mechanisms to determine mate choice. The signals oſten change as potential mates approach. These courtship sounds are modified advertisement calls which provide further opportunities for mate assessment. Acoustic signals can also be used intraspecifically to space individuals in the environment, as aggression signals, or as competitive signals to jam the signal of a neighbor. Eusocial insects use acoustic signals as warnings, to recruit a defensive response within the colony, and to recruit foragers to specific food sources.
Predator deterrence is another significant function of acoustic signals. The loud sounds produced by many insects (e.g., cicadas [Hemiptera]), can startle a potential predator giving the insect a chance to escape. The percussion sounds produced by acridid grasshoppers (Orthoptera) are thought to be defensive displays. Sound production has evolved specifically for this anti-predator function in click beetles (Coleoptera). The acoustic systems that use air movement in the Death’s Head moth (Lepidoptera), cockroaches (Blattodea) and grasshoppers (Orthoptera) also have anti-predator functions. Moths (Lepidoptera) have evolved a sound production system that is used to jam the ultrasonic signals of the bats that prey upon them. The arctiid moths emit ultrasonic pulses as bats approach which act to confuse the bat as to the exact location of its target. This gives the moth a chance to escape while the bat circles around for another attempt at capturing the insect. These pulses may also have an aposomatic function warning bats of a potentially distasteful meal.
Insects have also evolved specialized acoustic receptors as a means of avoiding predation. Bat-detectors have evolved independently in geometrid, noctuid and hawk moths (Lepidoptera), lacewings, (Neuroptera), praying mantises (Mantodea), beetles (Coleoptera), crickets, locusts and katydids (Orthoptera). These batdetectors provide an early warning system for the insects that a bat is nearby giving the insect a chance to escape before the bat can sense an insect in the vicinity.
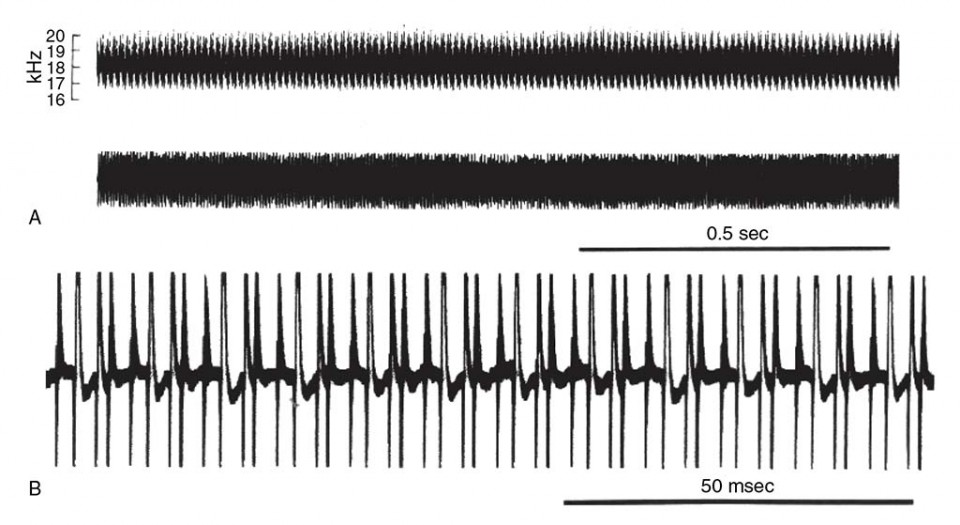
Figure 18 Acoustic signal produced by the cicada Beameria venosa (Uhler). (a) Sonogram of calling song illustrating energy distribution of the call. Middle trace is an oscillogram illustrating the temporal pattern of the call. (b) Expanded oscillogram to show individual sound pulses within the call.

Figure 19 Stridulatory apparatus of the cicada Tettigades undata Torres. The file (illustrated) is located on the mesothorax in this group of cicadas. The plectrum is located on the tegmina. Sound pulses are produced as the tegmina is rubbed over the series of mesothoracic ridges.
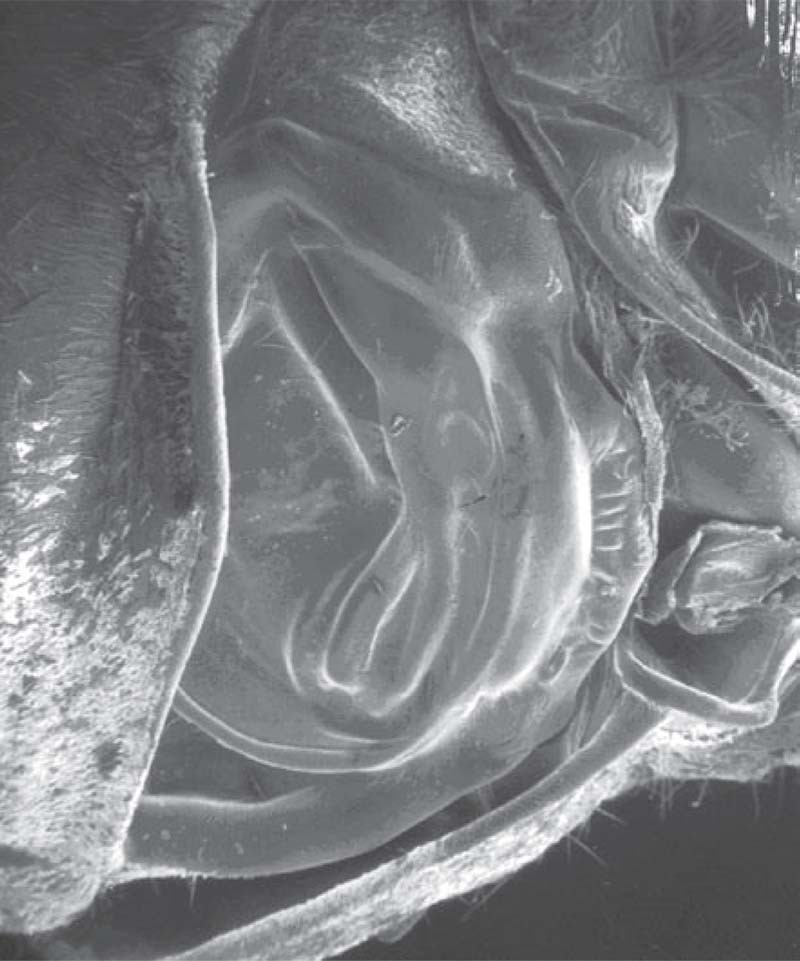
Figure 20 Tymbal organ of the cicada Beameria venosa (Uhler). The internally attached tymbal muscle buckles the tymbal to produce a sound pulse.

Acoustic Aposematism (Clicking) by Caterpillars


