Abdomen of Hexapods
The abdomen constitutes the caudal tagma in the hexapods and is usually larger than the other two, the head and the thorax. This region is also referred to as a visceral area because it houses the visceral organs. Its form can vary depending on the group, and even on the species. The maximum number of observed segments is 11, although certain authorities consider a twelfth segment that in fact corresponds to a telsonic caudal region. In general, the number of segments decreases from the preimaginal phases to the adult stage, especially in those holometabolous insects in which the last segments of the adults are formed from imaginal discs during pupation. In the groups considered most primitive, the number of abdominal segments is usually greater, as occurs in the Protura with 11 segments.
An exception is the Collembola, which only possess six. In addition, it is necessary to keep in mind that, in certain cases, the total number of visible segments does not coincide with what a particular individual actually possesses, since some segments remain “invisible” upon being telescoped, particularly those of the posterior region of the abdomen.
According to Bitsch, a generalized abdominal segment would be limited anteriorly by a presegmentary domain, separated from the segmentary domain proper (of greater size) by a suture that begins an internal crest named the costa or antecosta. This crest anteriorly delimits an acrotergite or precosta in the tergal part and a presternite in the sternal part. In this idealized model, the muscles would be inserted in successive antecostas. No known structure is homologous to the thoracic furca.
The presence of the gonopore (double in Ephemeroptera) in segments VIII and IX (in VII in the case of Ephemeroptera), and fundamentally of the external structures related to reproduction (the genitalia), produce important modifications in those segments. Considering the presence of these genitalia, three regions of the abdomen are recognized: an anterior (pregenital or visceral region that includes the first eight segments), median (genital region, eighth and ninth segments), and caudal regions (postgenital region, tenth and eleventh segments plus the telsonic region).
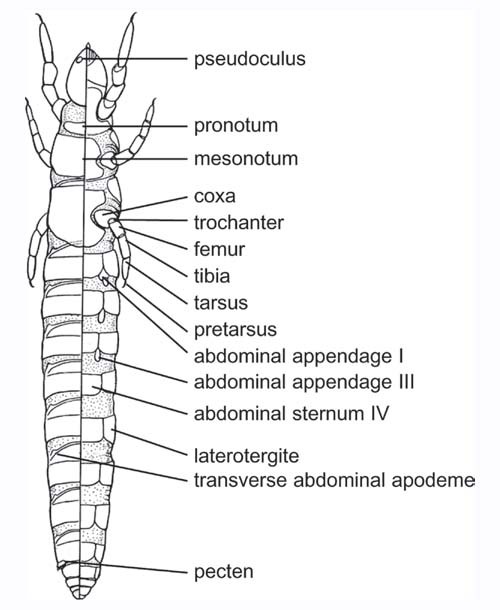
Abdomen of Hexapods, Diagram of a proturan (Protura) showing abdominal segments and appendages: dorsal view (left), ventral view (right).
The Pregenital Region
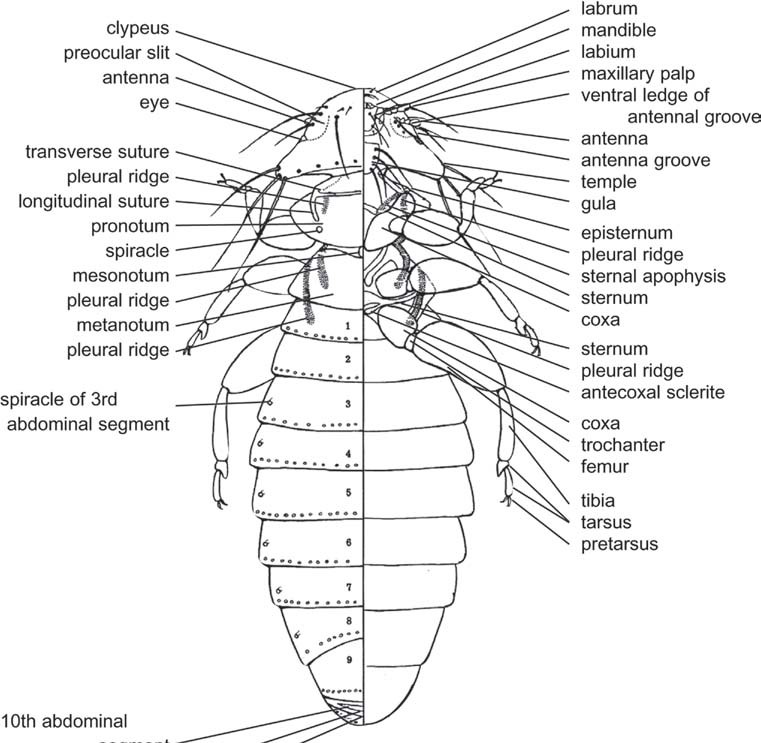
Diagram of chewing louse (Mallophaga) showing abdominal segments, including numbering of segments: dorsal view (left), ventral view (right).
In the most generalized condition, the first abdominal segments conserve their basic structure, being easily distinguished from the thoracic segments. Nevertheless, the most frequent condition is that which produces morphological modifications that affect the thoracic-abdominal union. These modifications usually consist of reductions that affect the sternal region and involve a greater or lesser desclerotization of different structures and their incorporation to the metathorax. In this sense, the case of the Hymenoptera, Apocrita stands out, in which a narrowing is produced between the second and the third abdominal segments, which incorporate the thorax and is named the propodeum. The rest of the abdominal segments are called the gaster or metasoma. The region formed by the propodeum and the thorax constitutes the mesosoma. The narrowing allows a great amplitude of movements of the metasoma, which permits stinging in the capture of prey in aculeates. In some groups, like Formicidae and Sphecidae (Aculeata), one or two segments of the metasoma form a narrower zone called the petiole.
In the pregenital region, several appendicular structures can be found. Thus, three pairs of highly modified appendages exist in Collembola. In Archaeognatha, very developed coxites are differentiated, above which are inserted styli in a median position and the exsertile vesicles in the most internal position.
The styli are elongated pieces, articulated in their base above the external face of the coxite. They are unisegmentary and lack muscles inserted in their base, often presenting an apical spine. Taking into account their position and their embryonic development, the styli are considered by the majority of authorities as vestigial appendages, and more concretely as reduced telepodites. The exsertile vesicles are considered internal coxal formations (internal coxalia of some authorities).
In Pterygota the abdominal appendages remain restricted to the larval forms (Lepidoptera and Hymenoptera, Tenthredinoidea), although rough appendicular pairs already exist in the polypodous type of embryos. These abdominal appendages are named “false legs” or “prolegs” and are retractile, conical and membranous projections, with a circular planta that bears a crown, usually with hooks, to adhere to the substrate.
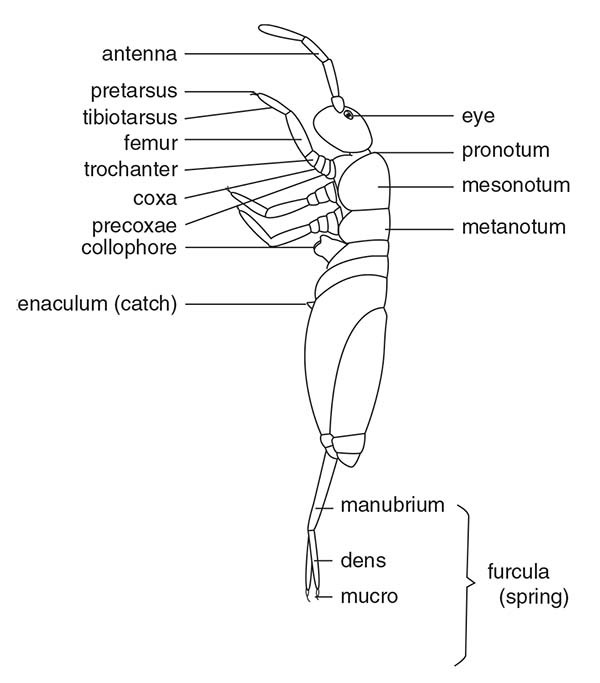
Diagram of springtail (Collembola) showing furcular appendage at tip of abdomen.
The Genital Region
The transformations that affect the eighth and ninth abdominal segments are a consequence of the development of special external structures that in the case of the male serve in the transfer of sperm, and in the case of the female allow for oviposition. These structures together are known by the name genitalia.
The origin of the genitalia is controversial, although the majority of authorities accept that, at least in part, it is of appendicular origin. In this sense, it is clear that in the Archaeognatha the eighth and ninth segments are basically similar in males and females and their structures are homologous to those already indicated for the pregenital segments. Taking into account this relationship, the genitalia of Archaeognatha are considered primitive, and therefore fundamental to interpret the genitalia of Pterygota.
In the eighth segment of the Archaeognatha, the basal part of the appendicular structure is named first gonocoxa or gonocoxite and bears the first gonostylus; in the ninth is found the second gonocoxa with its corresponding stylus. In both segments (at times in the eighth, always in the ninth) formations homologous to the exsertile vesicles appear, which are called gonapophyses (parameters in the males and gonapophysis proper in the females). The fundamental difference between both sexes lies in the presence in the males of a phallic structure.
The female genitalia in the Pterygota constitute the ovipositor. The gonocoxites are incorporated into the lateral wall of the genital segments in a complete manner in the eighth segment, forming the first valvifer. In the ninth segment the basal part is incorporated into the lateral wall, originating the second valvifer, while the rest is extended, forming the third pair of valves (dorsal or lateral valves of some authorities), which are not homologous in Archaeognatha. The other two pairs of valves are the ventral valves, corresponding to the eighth segment, and the internal valves, corresponding to the ninth segment. These two pairs of valves are homologous to the gonapophysis of Archaeognatha. In the case of the generalized type of ovipositor like that of Orthoptera, these three pairs of valves are linked through the length of their course, forming in their interior a canal for oviposition.
Among the sclerites that are situated in the base of the valves (in addition to the valvifers already mentioned) are found the intervalves (intervalvulae of the authorities) by way of elongated transverse formations, one in the base of the valves of the eighth segment and another in the base of the valves of the ninth segment. The typical ovipositor that was just described can experience modifications according to the functions that it carries out; one of the most drastic is found in Hymenoptera, Aculeata, where it is transformed into a sting that serves the females as an attacking organ, either to capture prey or as a defense. On the other hand, the process of oviposition can be carried out through other, different structures, as occurs in the females of certain Diptera. In that case, the last segments are retractile and the intersegmental zones are highly developed, in such a way that they can become telescoped, forming oviposition tubes; this type of “ovipositor” is named the ovicauda. Not being homologous to the genitalia, many authorities call it terminalia.
The masculine genitalia present great morphological variability, which together with their taxonomic importance, have been the object of an infinity of descriptions, many of them without truly anatomical criteria. This has originated the use of very varied terminologies that have done nothing but complicate its study and impede the establishment of homologies even in the same group, creating in this way a great nomenclatorial chaos.
In the males, in addition to the genitalia proper, other structures (processes, lobes, etc). exist that intervene in functions other than those strictly related to the transfer of sperm; among the most common is the grasping of the female during mating. It has already been mentioned that the majority of authorities consider that the interpretation of the genitalia of Pterygota should be made by homology with the basic condition that is found in Archaeognatha. In this group, the phallic complex is formed by a median organ, the phallus or penis, and a pair of segmented pieces named parameres, that in the case of maximum development can exist in the eighth and ninth segments. The parameres correspond to the gonapophysis of the females (although the term gonapophysis is utilized indistinctly for both sexes by some authorities).
Many morphological models have been proposed to describe the male genitalia of Pterygota. The most complete, since it gathers and discusses early data, is that proposed by Bitsch. According to this author, what together forms the copulatory organ (phallus or penis) and the structures associated with the parameres (considered in the sense expressed by the Archaeognatha) is named the phallic complex.
The aedeagus is a sclerotized tube, situated above a largely membranous phallobase, although in more complex cases the phallobase presents an internal fold that remains membranous (endotheca) while the external part is sclerotized (phallotheca or theca). The aedeagus presents an invagination that forms a more or less developed internal chamber (the endophallus), which communicates with the gonopore at its base and in the other extreme communicates with the exterior through the phallotreme. In counter-proposition to the endophallus, the part formed by the external walls of the phallobase and the aedeagus forms the ectophallus. The phallic complex can present variable development, even being able to cause the aedeagus to disappear, or on the contrary, increase in complexity, developing spines and other types of processes named flagellum, virga or pseudovirga over the internal walls of the endophallus. When the endotheca and the endophallus are evaginated, the genitalia are converted into authentic intromittent organs.
The primitive position of the male genitalia can be displaced through different types of turns; one of the most showy cases is that which occurs in some Hymenoptera, Symphyta that present the condition called strophandric, which is characterized by a 180° rotation of the genitalia. Rotations have also been observed in males of Diptera.
The postgenital region, as was mentioned in the beginning of this section, comprises the tenth and eleventh segments plus the telsonic region. The tenth segment has been detected in Protura, Diplura, Archaoegnatha, Thysanura (Zygentoma), Ephemeroptera, Plecoptera, and some Orthoptera. The morphology of this segment is basically similar to the pregenital segments, although with certain frequency it can form a ring when the tergum and sternum unite, or the sternal region can be membranous. In embryonic forms, a pair of appendicular outlines is seen above this segment. In certain holometabolous insects, structures of uncertain meaning appear, such as the socii of some Hymenoptera.
The eleventh segment is recognized in the majority of embryonic phases of hexapods. In Archaeognatha and Thysanura it forms an annular structure from whose dorsal part is differentiated a long and narrow process called filum terminale, while from the lateroventral position are differentiated the cerci that in the adults possess numerous divisions. In the Pterygotes, the eleventh segment is formed by the epiproct (tergal region) and the paraprocts (in the lateroventral position); in the more primitive groups exist cerci (whose length and number of divisions are variable) situated in the membranous zones that exist between the epiproct and the paraprocts. The telsonic, asegmentary region constitutes the perianal membrane or periproct.
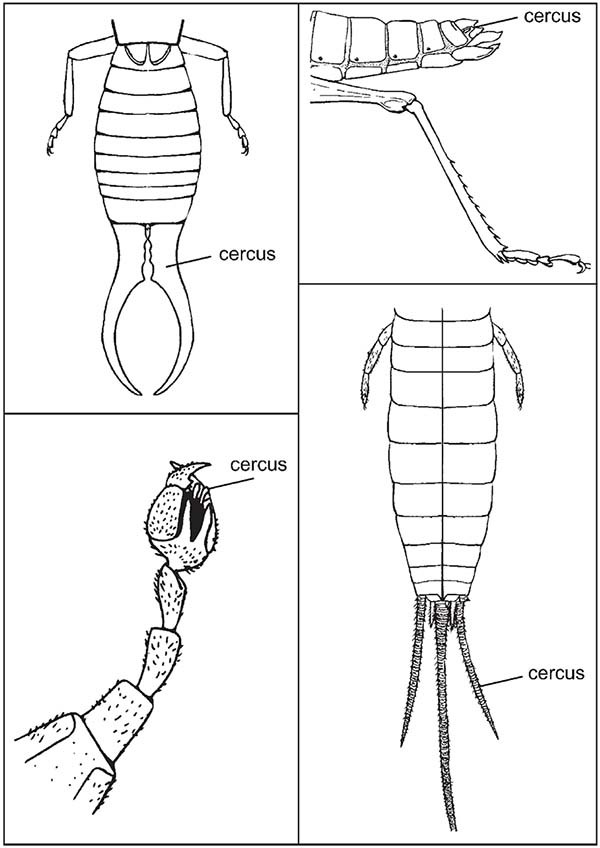
Comparative development of cerci on earwig (Dermaptera, top left); grasshopper (Orthoptera, top right); scorpion fly (Mecoptera, lower left); silverfish (Zygentoma).

Abbott’s Formula